

Drosophila are protected from Pseudomonas aeruginosa lethality by transgenic expression of paraoxonase-1
- PMID: 18704198
- PMCID: PMC2515384
- DOI: 10.1172/JCI35147
Drosophila are protected from Pseudomonas aeruginosa lethality by transgenic expression of paraoxonase-1
Abstract
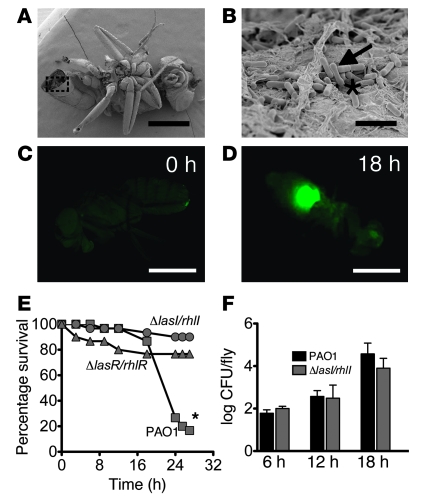
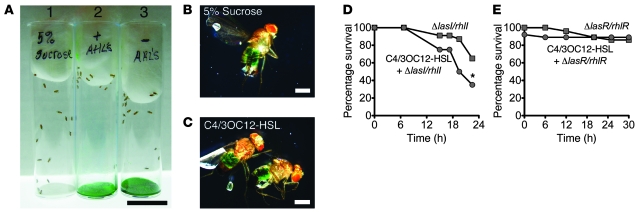
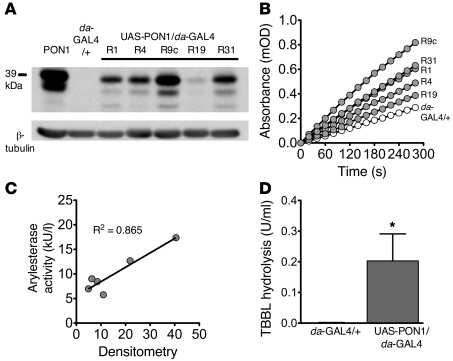
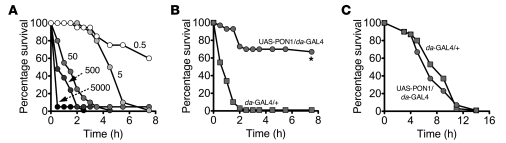
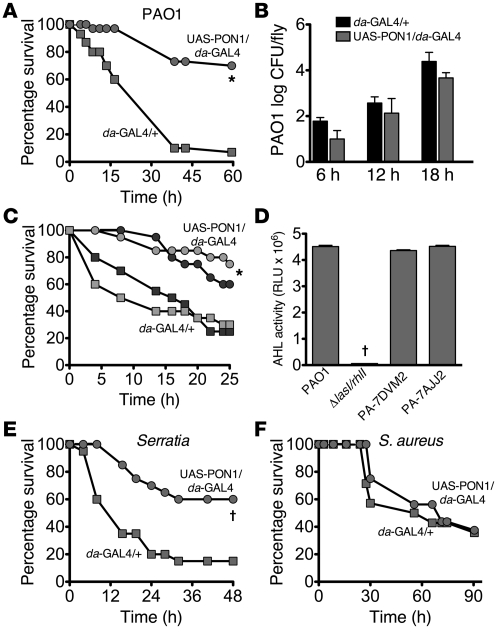
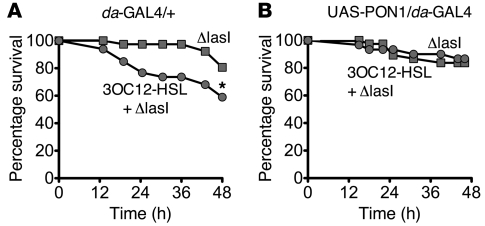
Pseudomonas aeruginosa uses quorum sensing, an interbacterial communication system, to regulate gene expression. The signaling molecule N-3-oxododecanoyl homoserine lactone (3OC12-HSL) is thought to play a central role in quorum sensing. Since 3OC12-HSL can be degraded by paraoxonase (PON) family members, we hypothesized that PONs regulate P. aeruginosa virulence in vivo. We chose Drosophila melanogaster as our model organism because it has been shown to be a tractable model for investigating host-pathogen interactions and lacks PONs. By using quorum-sensing-deficient P. aeruginosa, synthetic acyl-HSLs, and transgenic expression of human PON1, we investigated the role of 3OC12-HSL and PON1 on P. aeruginosa virulence. We found that P. aeruginosa virulence in flies was dependent upon 3OC12-HSL. PON1 transgenic flies expressed enzymatically active PON1 and thereby exhibited arylesterase activity and resistance to organophosphate toxicity. Moreover, PON1 flies were protected from P. aeruginosa lethality, and protection was dependent on the lactonase activity of PON1. Our findings show that PON1 can interfere with quorum sensing in vivo and provide insight into what we believe is a novel role for PON1 in the innate immune response to quorum-sensing-dependent pathogens. These results raise intriguing possibilities about human-pathogen interactions, including potential roles for PON1 as a modifier gene and for PON1 protein as a regulator of normal bacterial florae, a link between infection/inflammation and cardiovascular disease, and a potential therapeutic modality.
Figures

Similar articles
-
Dominant role of paraoxonases in inactivation of the Pseudomonas aeruginosa quorum-sensing signal N-(3-oxododecanoyl)-L-homoserine lactone.Infect Immun. 2008 Jun;76(6):2512-9. doi: 10.1128/IAI.01606-07. Epub 2008 Mar 17. Infect Immun. 2008. PMID: 18347034 Free PMC article.
-
Human and murine paraoxonase 1 are host modulators of Pseudomonas aeruginosa quorum-sensing.FEMS Microbiol Lett. 2005 Dec 1;253(1):29-37. doi: 10.1016/j.femsle.2005.09.023. Epub 2005 Oct 5. FEMS Microbiol Lett. 2005. PMID: 16260097
-
Paraoxonase 1, quorum sensing, and P. aeruginosa infection: a novel model.Adv Exp Med Biol. 2010;660:183-93. doi: 10.1007/978-1-60761-350-3_17. Adv Exp Med Biol. 2010. PMID: 20221881
-
Paraoxonase 1 status as a risk factor for disease or exposure.Adv Exp Med Biol. 2010;660:29-35. doi: 10.1007/978-1-60761-350-3_4. Adv Exp Med Biol. 2010. PMID: 20221868 Free PMC article. Review.
-
The roles of PON1 and PON2 in cardiovascular disease and innate immunity.Curr Opin Lipidol. 2009 Aug;20(4):288-92. doi: 10.1097/MOL.0b013e32832ca1ee. Curr Opin Lipidol. 2009. PMID: 19474728 Free PMC article. Review.
Cited by
-
Engineering human PON1 in an E. coli expression system.Adv Exp Med Biol. 2010;660:37-45. doi: 10.1007/978-1-60761-350-3_5. Adv Exp Med Biol. 2010. PMID: 20221869 Free PMC article.
-
Paraoxonases-1, -2 and -3: What are their functions?Chem Biol Interact. 2016 Nov 25;259(Pt B):51-62. doi: 10.1016/j.cbi.2016.05.036. Epub 2016 May 26. Chem Biol Interact. 2016. PMID: 27238723 Free PMC article.
-
Quorum Sensing Inhibition by Marine Bacteria.Mar Drugs. 2019 Jul 23;17(7):427. doi: 10.3390/md17070427. Mar Drugs. 2019. PMID: 31340463 Free PMC article. Review.
-
The Drosophila melanogaster host model.J Oral Microbiol. 2012;4. doi: 10.3402/jom.v4i0.10368. Epub 2012 Feb 21. J Oral Microbiol. 2012. PMID: 22368770 Free PMC article.
-
Thermostable Lactonases Inhibit Pseudomonas aeruginosa Biofilm: Effect In Vitro and in Drosophila melanogaster Model of Chronic Infection.Int J Mol Sci. 2023 Dec 1;24(23):17028. doi: 10.3390/ijms242317028. Int J Mol Sci. 2023. PMID: 38069351 Free PMC article.
References
-
- Gan K.N., Smolen A., Eckerson H.W., La Du B.N. Purification of human serum paraoxonase/arylesterase. Evidence for one esterase catalyzing both activities. Drug Metab. Dispos. 1991;19:100–106. - PubMed
-
- Durrington P.N., Mackness B., Mackness M.I. Paraoxonase and atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2001;21:473–480. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
