

Hepatocyte nuclear factor 4alpha is implicated in endoplasmic reticulum stress-induced acute phase response by regulating expression of cyclic adenosine monophosphate responsive element binding protein H
- PMID: 18704925
- PMCID: PMC2717709
- DOI: 10.1002/hep.22439
Hepatocyte nuclear factor 4alpha is implicated in endoplasmic reticulum stress-induced acute phase response by regulating expression of cyclic adenosine monophosphate responsive element binding protein H
Abstract
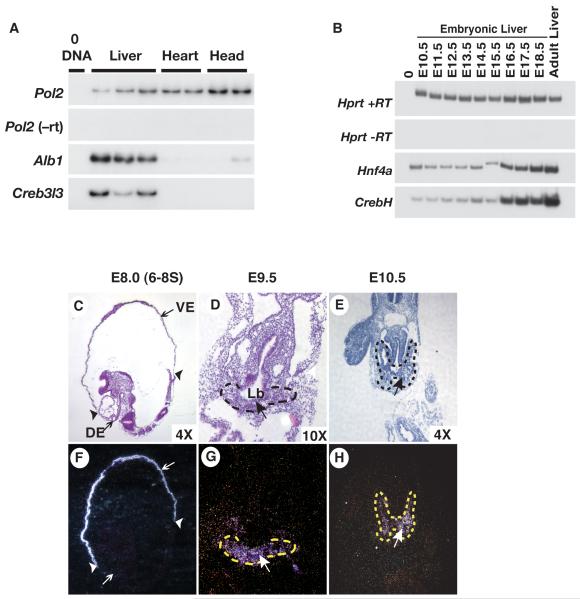
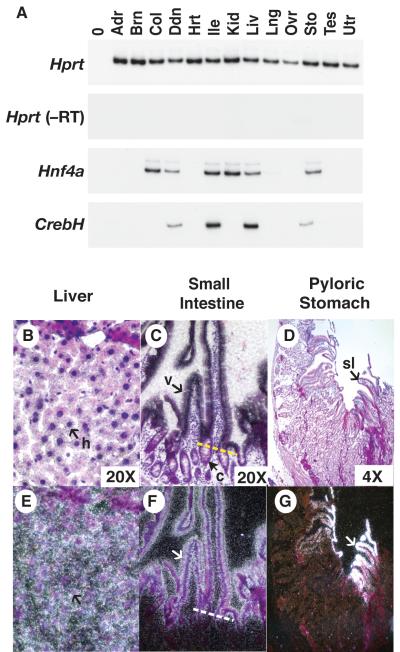
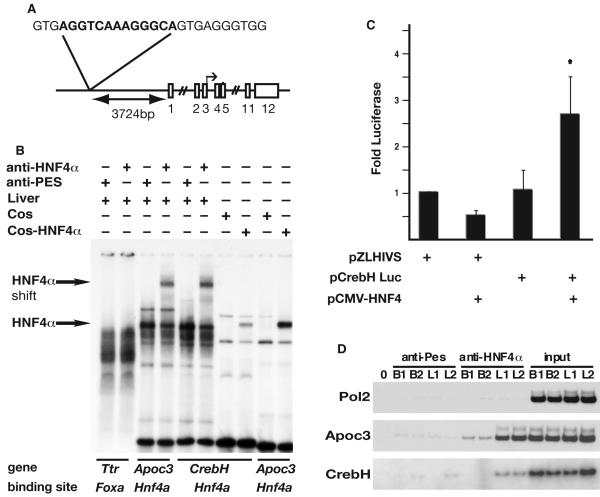
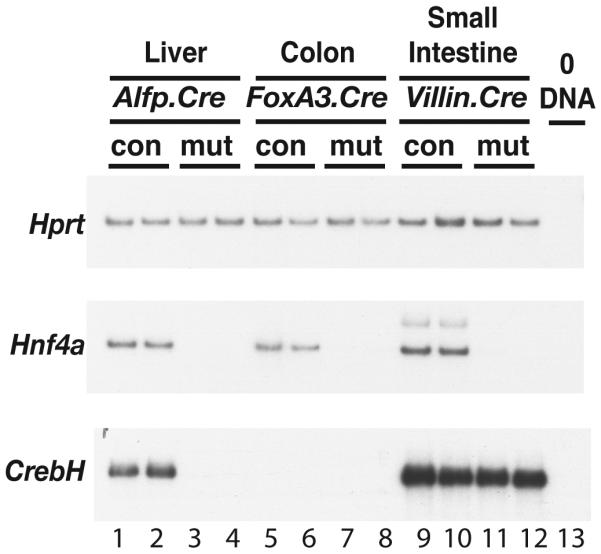
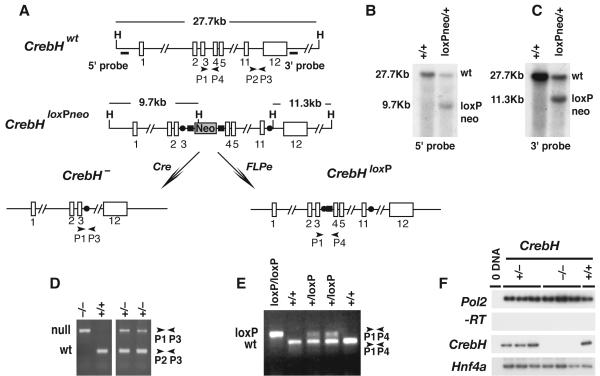
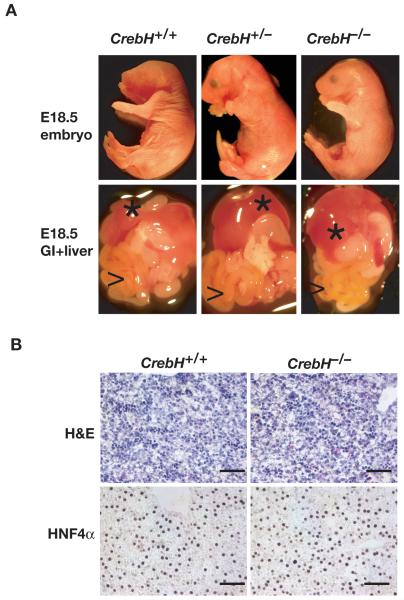
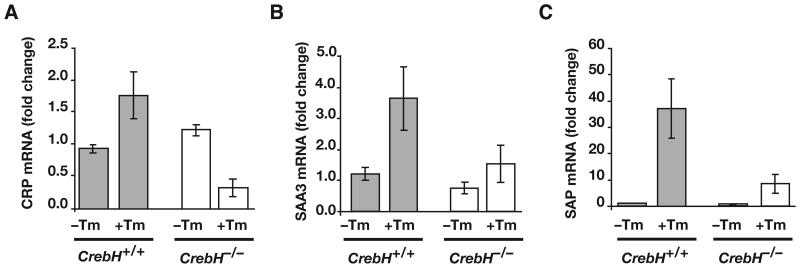
Loss of the nuclear hormone receptor hepatocyte nuclear factor 4alpha (HNF4alpha) in hepatocytes results in a complex pleiotropic phenotype that includes a block in hepatocyte differentiation and a severe disruption to liver function. Recent analyses have shown that hepatic gene expression is severely affected by the absence of HNF4alpha, with expression of 567 genes reduced by > or =2.5-fold (P < or = 0.05) in Hnf4alpha(-/-) fetal livers. Although many of these genes are direct targets, HNF4alpha has also been shown to regulate expression of other liver transcription factors, and this raises the possibility that the dependence on HNF4alpha for normal expression of some genes may be indirect. We postulated that the identification of transcription factors whose expression is regulated by HNF4alpha might reveal roles for HNF4alpha in controlling hepatic functions that were not previously appreciated. Here we identify cyclic adenosine monophosphate responsive element binding protein H (CrebH) as a transcription factor whose messenger RNA can be identified in both the embryonic mouse liver and adult mouse liver and whose expression is dependent on HNF4alpha. Analyses of genomic DNA revealed an HNF4alpha binding site upstream of the CrebH coding sequence that was occupied by HNF4alpha in fetal livers and facilitated transcriptional activation of a reporter gene in transient transfection analyses. Although CrebH is highly expressed during hepatogenesis, CrebH(-/-) mice were viable and healthy and displayed no overt defects in liver formation. However, upon treatment with tunicamycin, which induces an endoplasmic reticulum (ER)-stress response, CrebH(-/-) mice displayed reduced expression of acute phase response proteins.
Conclusion: These data implicate HNF4alpha in having a role in controlling the acute phase response of the liver induced by ER stress by regulating expression of CrebH.
Figures
References
-
- Parviz F, Matullo C, Garrison WD, Savatski L, Adamson JW, Ning G, Kaestner KH, et al. Hepatocyte nuclear factor 4alpha controls the development of a hepatic epithelium and liver morphogenesis. Nat Genet. 2003;34:292–296. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials