

Citrus tristeza virus: a pathogen that changed the course of the citrus industry
- PMID: 18705856
- PMCID: PMC6640355
- DOI: 10.1111/j.1364-3703.2007.00455.x
Citrus tristeza virus: a pathogen that changed the course of the citrus industry
Abstract
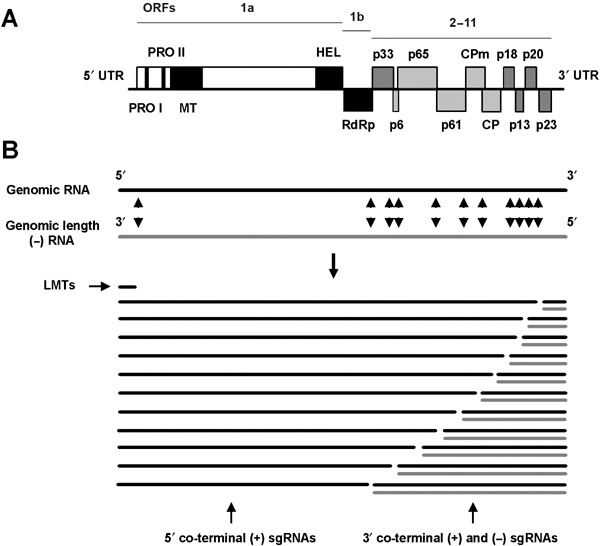
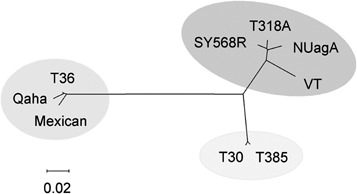
Citrus tristeza virus (CTV) (genus Closterovirus, family Closteroviridae) is the causal agent of devastating epidemics that changed the course of the citrus industry. Adapted to replicate in phloem cells of a few species within the family Rutaceae and to transmission by a few aphid species, CTV and citrus probably coevolved for centuries at the site of origin of citrus plants. CTV dispersal to other regions and its interaction with new scion varieties and rootstock combinations resulted in three distinct syndromes named tristeza, stem pitting and seedling yellows. The first, inciting decline of varieties propagated on sour orange, has forced the rebuilding of many citrus industries using tristeza-tolerant rootstocks. The second, inducing stunting, stem pitting and low bearing of some varieties, causes economic losses in an increasing number of countries. The third is usually observed by biological indexing, but rarely in the field. CTV polar virions are composed of two capsid proteins and a single-stranded, positive-sense genomic RNA (gRNA) of approximately 20 kb, containing 12 open reading frames (ORFs) and two untranslated regions (UTRs). ORFs 1a and 1b, encoding proteins of the replicase complex, are directly translated from the gRNA, and together with the 5' and 3'UTRs are the only regions required for RNA replication. The remaining ORFs, expressed via 3'-coterminal subgenomic RNAs, encode proteins required for virion assembly and movement (p6, p65, p61, p27 and p25), asymmetrical accumulation of positive and negative strands during RNA replication (p23), or suppression of post-transcriptional gene silencing (p25, p20 and p23), with the role of proteins p33, p18 and p13 as yet unknown. Analysis of genetic variation in CTV isolates revealed (1) conservation of genomes in distant geographical regions, with a limited repertoire of genotypes, (2) uneven distribution of variation along the gRNA, (3) frequent recombination events and (4) different selection pressures shaping CTV populations. Measures to control CTV damage include quarantine and budwood certification programmes, elimination of infected trees, use of tristeza-tolerant rootstocks, or cross protection with mild isolates, depending on CTV incidence and on the virus strains and host varieties predominant in each region. Incorporating resistance genes into commercial varieties by conventional breeding is presently unfeasible, whereas incorporation of pathogen-derived resistance by plant transformation has yielded variable results, indicating that the CTV-citrus interaction may be more specific and complex than initially thought. A deep understanding of the interactions between viral proteins and host and vector factors will be necessary to develop reliable and sound control measures.
Figures

References
-
- Albiach‐Martí, M.R. , Guerri, J. , Cambra, M. , Garnsey, S.M. and Moreno, P. (2000a) Differentiation of citrus tristeza virus isolates by serological analysis of the p25 coat protein peptide maps. J. Virol. Methods, 88, 25–34. - PubMed
-
- Albiach‐Martí, M.R. , Guerri, J. , Hermoso de Mendoza, A. , Laigret, F. , Ballester‐Olmos, J.F. and Moreno, P. (2000b) Aphid transmission alters the genomic and defective RNA populations of citrus tristeza virus. Phytopathology, 90, 134–138. - PubMed
-
- Albiach‐Martí, M.R. , Mawassi, M. , Gowda, S. , Satyanayanana, T. , Hilf, M.E. , Shanker, S. , Almira, E.C. , Vives, M.C. , López, C. , Guerri, J. , Flores, R. , Moreno, P. , Garnsey, S.M. and Dawson, W.O. (2000c) Sequences of Citrus tristeza virus separated in time and space are essentially identical. J. Virol. 74, 6856–6865. - PMC - PubMed
-
- Albiach‐Martí, M.R. , Grosser, J.W. , Gowda, S. , Mawassi, M. , Satyanarayana, T. , Garnsey, S.M. and Dawson, W.O. (2004) Citrus tristeza virus replicates and forms infectious virions in protoplast of resistant citrus relatives. Mol. Breeding, 14, 117–128.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
