

Phytoplasmas: bacteria that manipulate plants and insects
- PMID: 18705857
- PMCID: PMC6640453
- DOI: 10.1111/j.1364-3703.2008.00472.x
Phytoplasmas: bacteria that manipulate plants and insects
Abstract
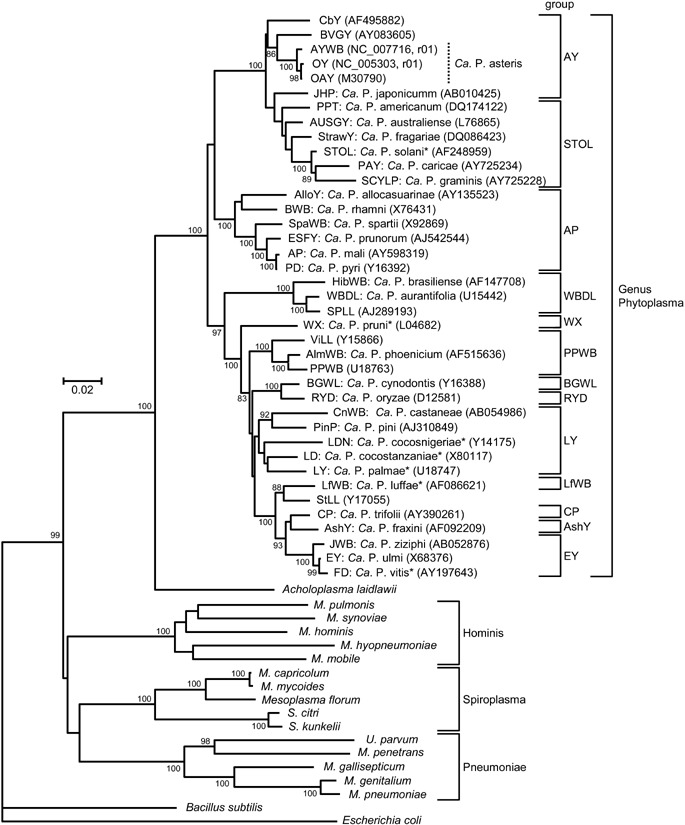
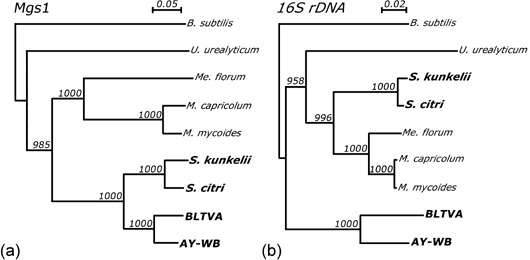
Taxonomy: Superkingdom Prokaryota; Kingdom Monera; Domain Bacteria; Phylum Firmicutes (low-G+C, Gram-positive eubacteria); Class Mollicutes; Candidatus (Ca.) genus Phytoplasma.
Host range: Ca. Phytoplasma comprises approximately 30 distinct clades based on 16S rRNA gene sequence analyses of approximately 200 phytoplasmas. Phytoplasmas are mostly dependent on insect transmission for their spread and survival. The phytoplasma life cycle involves replication in insects and plants. They infect the insect but are phloem-limited in plants. Members of Ca. Phytoplasma asteris (16SrI group phytoplasmas) are found in 80 monocot and dicot plant species in most parts of the world. Experimentally, they can be transmitted by approximately 30, frequently polyphagous insect species, to 200 diverse plant species.
Disease symptoms: In plants, phytoplasmas induce symptoms that suggest interference with plant development. Typical symptoms include: witches' broom (clustering of branches) of developing tissues; phyllody (retrograde metamorphosis of the floral organs to the condition of leaves); virescence (green coloration of non-green flower parts); bolting (growth of elongated stalks); formation of bunchy fibrous secondary roots; reddening of leaves and stems; generalized yellowing, decline and stunting of plants; and phloem necrosis. Phytoplasmas can be pathogenic to some insect hosts, but generally do not negatively affect the fitness of their major insect vector(s). In fact, phytoplasmas can increase fecundity and survival of insect vectors, and may influence flight behaviour and plant host preference of their insect hosts.
Disease control: The most common practices are the spraying of various insecticides to control insect vectors, and removal of symptomatic plants. Phytoplasma-resistant cultivars are not available for the vast majority of affected crops.
Figures






References
-
- Adam, S.A. , Adam, E.J. , Chi, N.C. and Visser, G.D. (1995) Cytoplasmic factors in NLS‐mediated targeting to the nuclear pore complex. Cold Spring Harb. Symp. Quant. Biol. 60, 687–694. - PubMed
-
- Ammar, E.‐D. and Hogenhout, S. A. (2006) Mollicutes associated with arthropods and plants In: Insect Symbiosis, Vol. 2 (Kostas B. and Miller T., eds), pp. 97–118. CRC Press, Taylor and Francis Group, Boca Raton, FL, USA.
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
