

Natural variation reveals key amino acids in a downy mildew effector that alters recognition specificity by an Arabidopsis resistance gene
- PMID: 18705864
- PMCID: PMC6640421
- DOI: 10.1111/j.1364-3703.2008.00481.x
Natural variation reveals key amino acids in a downy mildew effector that alters recognition specificity by an Arabidopsis resistance gene
Abstract
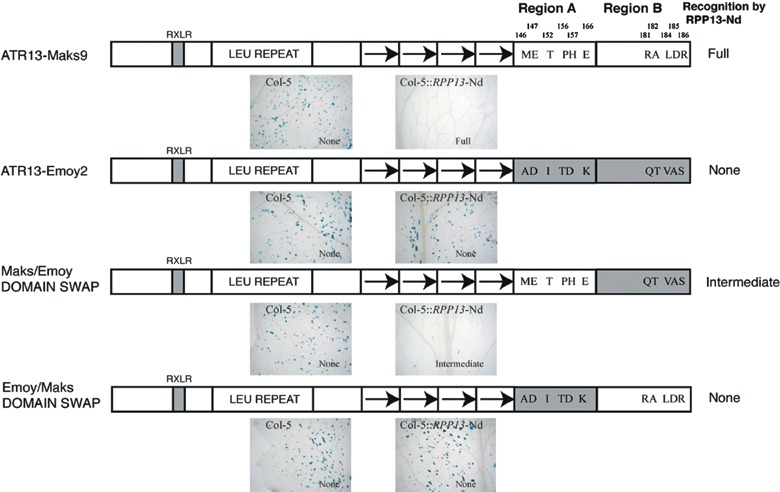
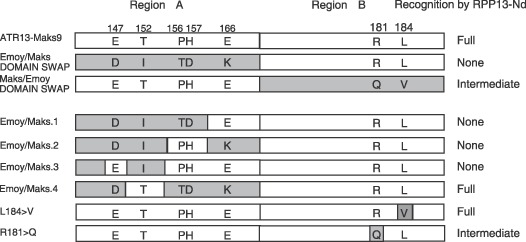
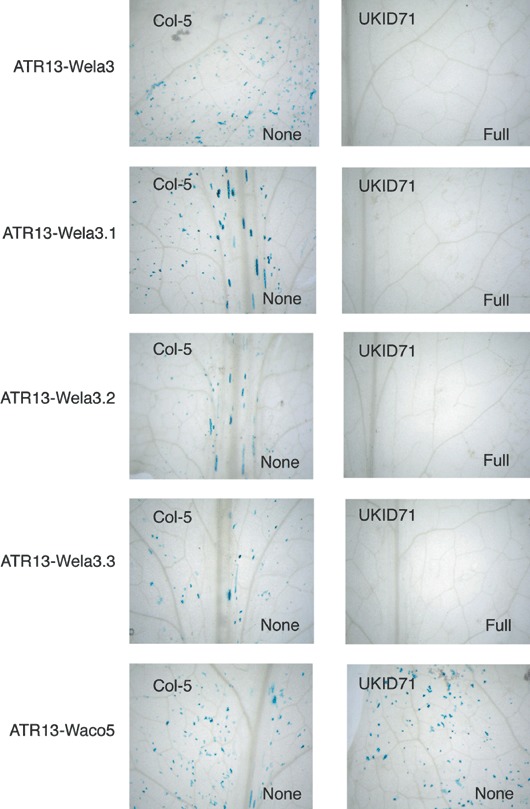
RPP13, a member of the cytoplasmic class of disease resistance genes, encodes one of the most variable Arabidopsis proteins so far identified. This variability is matched in ATR13, the protein from the oomycete downy mildew pathogen Hyaloperonospora parasitica recognized by RPP13, suggesting that these proteins are involved in tight reciprocal coevolution. ATR13 exhibits five domains: an N-terminal signal peptide, an RXLR motif, a heptad leucine/isoleucine repeat, an 11-amino-acid repeated sequence and a C-terminal domain. We show that the conserved RXLR-containing domain is dispensable for ATR13-mediated recognition, consistent with its role in transport into the plant cytoplasm. Sequencing ATR13 from 16 isolates of H. parasitica revealed high levels of amino acid diversity across the entire protein. The leucines/isoleucines of the heptad leucine repeat were conserved, and mutation of particular leucine or isoleucine residues altered recognition by RPP13. Natural variation has not exploited this route to detection avoidance, suggesting a key role of this domain in pathogenicity. The extensive variation in the 11-amino-acid repeat units did not affect RPP13 recognition. Domain swap analysis showed that recognition specificity lay in the C-terminal domain of ATR13. Variation analyses combined with functional assays allowed the identification of four amino acid positions that may play a role in recognition specificity. Site-directed mutagenesis confirmed that a threonine residue is absolutely required for RPP13 recognition and that recognition can be modulated by the presence of either an arginine or glutamic acid at other sites. Mutations in these three amino acids had no effect on the interaction of ATR13 with a resistance gene unlinked to RPP13, consistent with their critical role in determining RPP13-Nd recognition specificity.
Figures



References
-
- Abramovitch, R.B. and Martin, G.B. (2004) Strategies used by bacterial pathogens to suppress plant defenses. Curr. Opin. Plant. Biol. 7, 356–364. - PubMed
-
- Van Den Ackerveken, G. , Marois, E. and Bonas, U. (1996) Recognition of the bacterial avirulence protein AvrBs3 occurs inside the host plant cell. Cell, 87, 1307–1316. - PubMed
-
- Allen, R.L. , Bittner‐Eddy, P.D. , Grenville‐Briggs, L.J. , Meitz, J.C. , Rehmany, A.P. , Rose, L.E. and Beynon, J.L. (2004). Host–parasite coevolutionary conflict between Arabidopsis and downy mildew. Science, 306, 1957–1960. - PubMed
-
- Armstrong, M.R. , Whisson, S.C. , Pritchard, L. , Bos, J.I.B. , Venter, E. , Avrova, A.O. , Rehmany, A.P. , Bohme, U. , Brooks, K. , Cherevach, I. , Hamlin, N. , White, B. , Frasers, A. , Lord, A. , Quail, M.A. , Churcher, C. , Hall, N. , Berriman, M. , Huang, S. , Kamoun, S. , Beynon, J.L. and Birch, P.R.J. (2005). An ancestral oomycete locus contains late blight avirulence gene Avr3a, encoding a protein that is recognized in the host cytoplasm. Proc. Natl. Acad. Sci. USA, 102, 7766–7771. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
