

Review
doi: 10.1016/j.resp.2008.07.014.
The chemical neuroanatomy of breathing
Affiliations
- PMID: 18706532
- PMCID: PMC2701569
- DOI: 10.1016/j.resp.2008.07.014
Item in Clipboard
Review
The chemical neuroanatomy of breathing
Respir Physiol Neurobiol.
.
Abstract
The chemical neuroanatomy of breathing must ultimately encompass all the various neuronal elements physiologically identified in brainstem respiratory circuits and their apparent aggregation into "compartments" within the medulla and pons. These functionally defined respiratory compartments in the brainstem provide the major source of input to cranial motoneurons controlling the airways, and to spinal motoneurons activating inspiratory and expiratory pump muscles. This review provides an overview of the neuroanatomy of the major compartments comprising brainstem respiratory circuits, and a synopsis of the transmitters used by their constituent respiratory neurons.
Figures
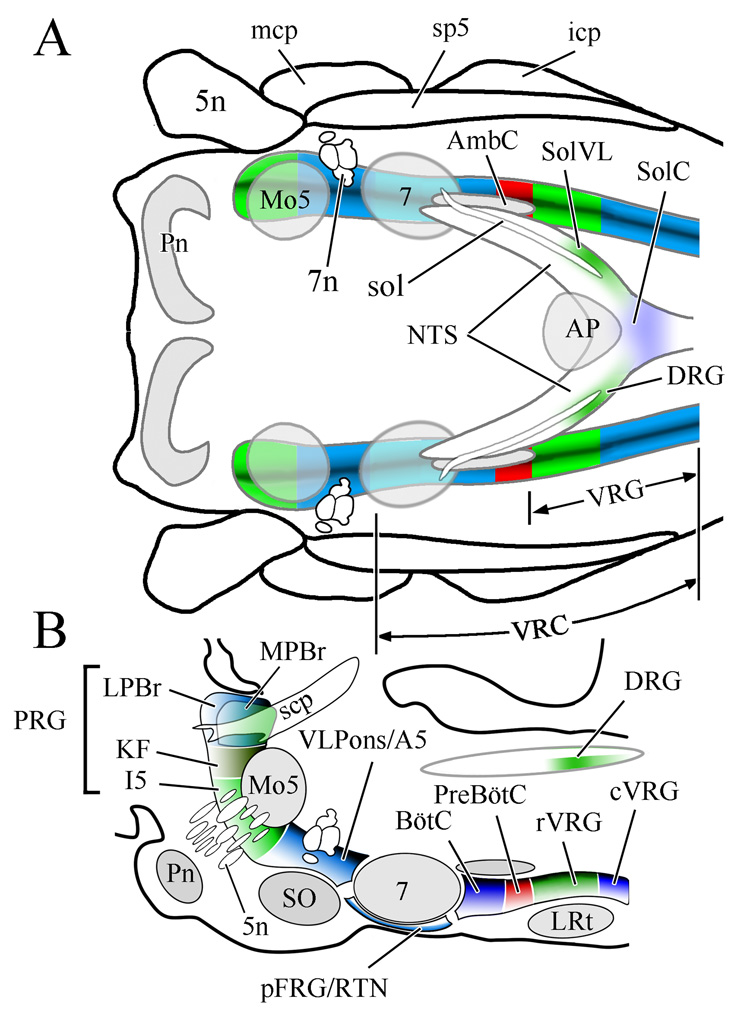
Respiratory related regions of the rhombencephalon of the rat shown in horizontal (A) and sagittal (B) views. Note that respiratory related regions comprise a nearly continuous column in the lateral rhombencephalon. The boundaries depicted between the various brainstem compartments reflect functional distinctions between adjacent regions relative to their impact on breathing. Reproduced from McCrimmon et al., 2008 with permission; B is redrawn after Fig 1B in Alheid et al., 2004). Abbreviations, including subsequent figures: 5n, trigeminal nerve; 7, facial nucleus; 7n, facial nerve; A5, A5 noradrenergic neuronal group; AmbC, compact part of nucleus ambiguus; AP, area postrema; BötC, Bötzinger complex; cVRG, caudal division of ventral respiratory group; DRG, dorsal respiratory group; I5, intertrigeminal area; icp, inferior cerebellar peduncle; KF, Kölliker-Fuse nucleus; LPBr, lateral parabrachial region; LRt, lateral reticular nucleus; mcp, medial cerebellar peduncle; Mo5, motor nucleus of the trigeminal nerve; MPBr, medial parabrachial region; NTS, nucleus of the solitary tract; pFRG, parafacial respiratory group; Pn, basilar pontine nuclei; preBötC, preBötzinger complex; PRG, pontine respiratory group; RTN, retrotrapezoid nucleus; rVRG, rostral division of ventral respiratory group; scp, superior cerebellar peduncle; SO, superior olive; sol, solitary tract; SolC, commissural subdivision of the nucleus of the solitary tract; SolVL, ventrolateral subdivision of the nucleus of the solitary tract; sp5, spinal trigeminal tract; vlPons, ventrolateral pontine region; VRC, ventral respiratory column of the medulla; VRG, ventral respiratory group.

Discharge patterns of representative types of VRC respiratory neurons in the rat, depicted over three breaths. The abbreviations, glu, gly, and GABA identify the excitatory or inhibitory fast amino acid transmitters (glutamate, glycine, GABA) used by these cells where they have been experimentally established (see text). In instances where the excitatory or inhibitory nature of a particular cell type has been inferred, but where the transmitter has not been established the cells are marked with + or − signs. In the chart higher action potential frequencies are represented by denser color coding in the temporal firing pattern. Note that most current models account for three phases in the respiratory cycle: Inspiration (I-red), early expiration (E1-light blue), late expiration (E2-dark blue). This is supported by the observation of subsets of expiratory neurons whose active phase of firing only occurs during the early or late phase of expiration under normal relaxed breathing (eupnea). Neuron types include those with decrementing (Dec), constant (Con), or augmenting firing patterns (Aug). Inhibitory E-Dec neurons are included at two levels to reflect the presence of similar inhibitory expiratory neurons rostrally and caudally in the VRC (in the BötC and in the cVRG, respectively). A complete taxonomy of the various neuronal types suggested in different labs, is not encompassed in this single figure, neither have we included alternative terminologies used to designate the neuron types depicted. It is also acknowledged that under various environmental regimens the pattern of firing for individual neuronal types appears to be mutable. Finally, the excitatory and inhibitory nature of the neuronal types depicted represents cells examined in the VRC. While comparable inspiratory and expiratory firing patterns are observed in dorsolateral pontine and NTS neurons, the excitatory and inhibitory transmitters used by these neurons are generally not well defined.

Comparative distribution and functions of brainstem respiratory neurons. A: Outline of respiratory compartments in the rat VRC. The shaded regions indicate an approximate division of the VRC into a rostral part contributing to rhythm and pattern generation (pink), while more caudal regions appear to contribute only to the pattern of respiratory activity (green). B: Relative distribution of expiratory and inspiratory neuron subtypes in the VRC. Data adapted from plots of propriobulbar neurons recorded in the rat by Ezure et al. (1988; E-Aug, E-Dec, I-Aug, I-Dec, I-Con) & by Sun et al. (1998; E-I and I-E) and normalized to the peak number of expiratory neurons in BötC. Note that the distribution maxima for particular neuron subtypes corresponds to particular VRC compartments, but that the sharp borders shown in the cartoon in A do not adequately represent the actual blending of different cell distributions along the rostral caudal length of the VRC. C. Effect on expiratory duration (Te ) of the excitatory amino acid DL-homocysteic acid (DLH) micro-stimulation of the BötC, preBötC, and rVRG. Redrawn with permission after Monnier et al., 2003. Note, stimulation in preBötC caused tachypnea (shortened Te ) while stimulation in either the BötC or the anterior part of rVRG produced bradypnea (lengthened Te ). Chemical stimulation in posterior portions of the rVRG does not affect respiratory rate (Wang et al., 2002; Monnier et al., 2003).

NK1 receptors in the rostral VRC of the mouse. Pseudocolored sagittal section through the rhombencephalon of the mouse with immunolabeled NK1 receptors (cyan). Note the dense labeling at the level of the preBötC and also at the ventral and caudal portions of the facial nucleus. Nevertheless, in comparison to areas such as the superior olive (SO) and lateral reticular nucleus (LRt), which are essentially unstained for NK1 receptors, few areas of the VRC (i.e. the RTN, BötC, preBötC, or rVRG) can be considered devoid of NK1 receptors. Following immunolabeling with diaminobenzidine as the chromogen, the section was subsequently counterstained with ethidium bromide (red) as a fluorescent Nissl stain. Grayscale images of the NK1 receptors were inverted with respect to black and white and copied to the green and blue channels of an RGB image. A grayscale image of the Nissl staining was copied to the red channel of the same image to produce the final composite image.
References
-
- Alheid GF, Gray PA, Jiang MC, Feldman JL, McCrimmon DR. Parvalbumin in respiratory neurons of the ventrolateral medulla of the adult rat. J Neurocytol. 2002;31:693–717. - PubMed
-
- Alheid GF, Milsom WK, McCrimmon DR. Pontine influences on breathing: an overview. Respir Physiol Neurobiol. 2004;143:105–114. - PubMed
-
- Amiel J, Laudier B, Attie-Bitach T, Trang H, de Pontual L, Gener B, Trochet D, Etchevers H, Ray P, Simonneau M, Vekemans M, Munnich A, Gaultier C, Lyonnet S. Polyalanine expansion and frameshift mutations of the paired-like homeobox gene PHOX2B in congenital central hypoventilation syndrome. Nat Genet. 2003;33:459–461. - PubMed
-
- Averill DB, Cameron WE, Berger AJ. Neural elements subserving pulmonary stretch receptor-mediated facilitation of phrenic motoneurons. Brain Res. 1985;346:378–382. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
