

Partial infraorbital nerve ligation as a model of trigeminal nerve injury in the mouse: behavioral, neural, and glial reactions
- PMID: 18708302
- PMCID: PMC2632609
- DOI: 10.1016/j.jpain.2008.06.006
Partial infraorbital nerve ligation as a model of trigeminal nerve injury in the mouse: behavioral, neural, and glial reactions
Abstract
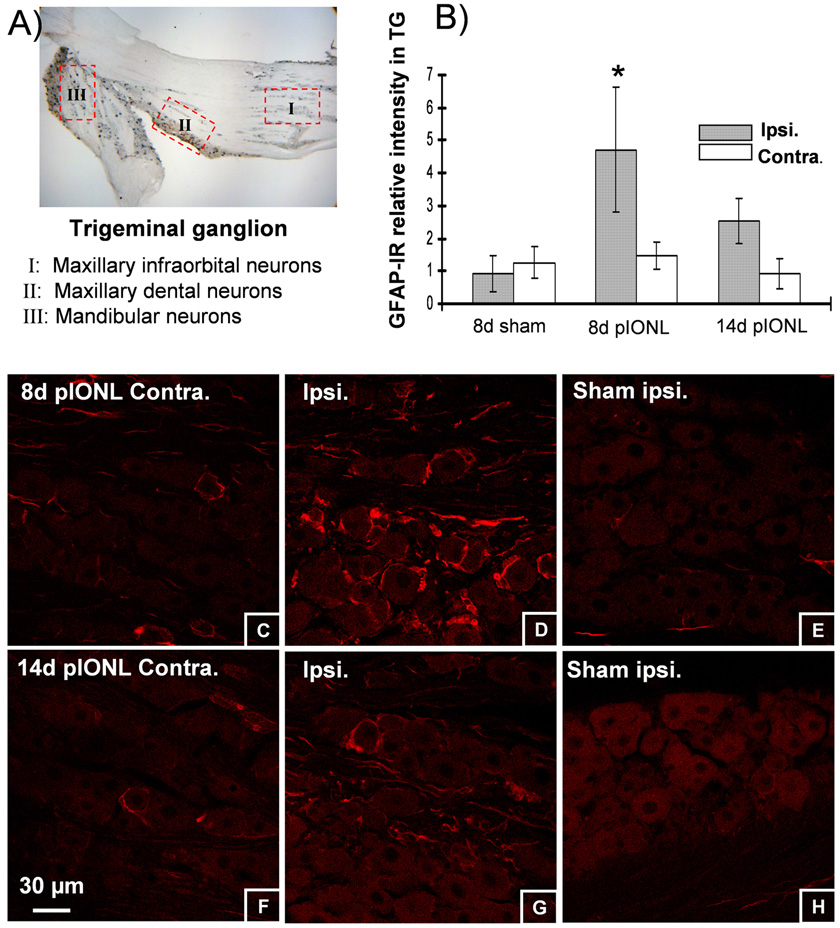
Trigeminal nerve damage often leads to chronic pain syndromes including trigeminal neuralgia, a severely debilitating chronic orofacial pain syndrome. Options for treatment of neuropathic pain are limited in effectiveness and new approaches based on a better understanding of the underlying pathologies are required. Partial ligation has been shown to effectively mimic many of the qualities of human neuropathic pain syndromes. We have devised a mouse model of trigeminal neuralgia using a partial infraorbital nerve ligation (pIONL) that induces persistent pain behaviors and morphological changes in the brainstem. We found that the pIONL effectively induced mechanical allodynia lasting for more than 3 weeks. Cell proliferation (bromodeoxyuridine), activation of astrocytes and microglia in the ipsilateral caudal medulla, and persistent satellite cell reaction in the ipsilateral ganglion were observed. Neurochemical markers calcitonin gene-related peptide, substance P were decreased in medullary dorsal horn ipsilateral to the injury side, whereas substance P receptor NK1 expression was increased after 8 days. Nerve injury marker ATF3 was markedly increased in ipsilateral trigeminal ganglion neurons at 8 days after pIONL. The data indicate that partial trigeminal injury in mice produces many persistent anatomical changes in neuropathic pain, as well as mechanical allodynia.
Perspective: This study describes the development of a new mouse model of trigeminal neuropathic pain. Our goal is to devise better treatments of trigeminal pain, and this will be facilitated by characterization of the underlying cellular and molecular neuropathological mechanisms in genetically designed mice.
Figures







References
-
- Abbadie C, Brown JL, Mantyh PW, Basbaum AI. Spinal cord substance P receptor immunoreactivity increases in both inflammatory and nerve injumry models of persistent pain. Neuroscience. 1996;70:201–209. - PubMed
-
- Anderson LC, von Bartheld CS, Byers MR. NGF depletion reduces ipsilateral and contrlateral trigeminal satellite cell reactions after inferior alveolar nerve injury in adult rats. Exp Neurol. 1998;150:312–320. - PubMed
-
- Anderson LC, Vakoula A, Veinote R. Inflammatory hypersensitivity in a rat model of trigeminal neuropathic pain. Arch Oral Biol. 2003;48:161–169. - PubMed
-
- Averill S, Michael GJ, Shortland PJ, Leavesley RC, King VR, Bradbury EJ, McMahon SB, Priestley JV. NGF and GDNF ameliorate the increase in ATF3 expression which occurs in dorsal rootganglion cells in response to peripheral nerve injury. Eur. J. Neurosci. 2004;19:1437–1445. - PubMed
-
- Bennett GJ, Xie YK. A peripheral mononeuropathy in rat that produces disorders of pain sensation like those seen in man. Pain. 1988;33:87–107. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
