

Selective distribution of protein kinase A regulatory subunit RII{alpha} in rodent gliomas
- PMID: 18708342
- PMCID: PMC2719010
- DOI: 10.1215/15228517-2008-054
Selective distribution of protein kinase A regulatory subunit RII{alpha} in rodent gliomas
Abstract
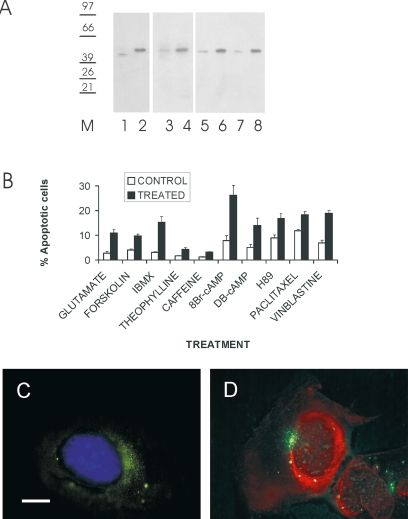
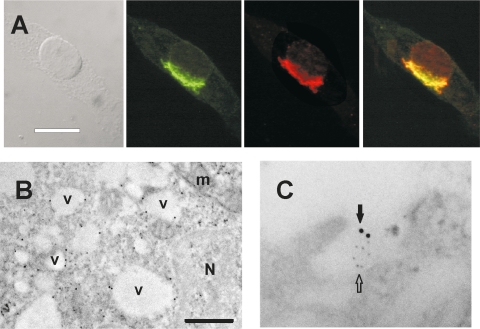
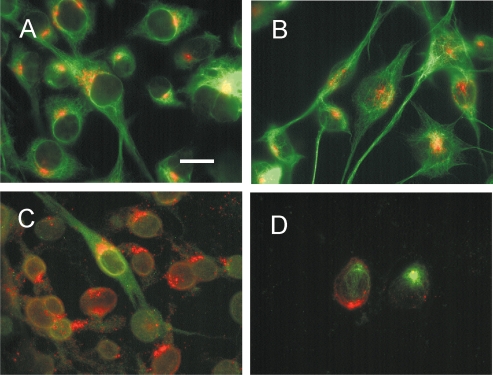
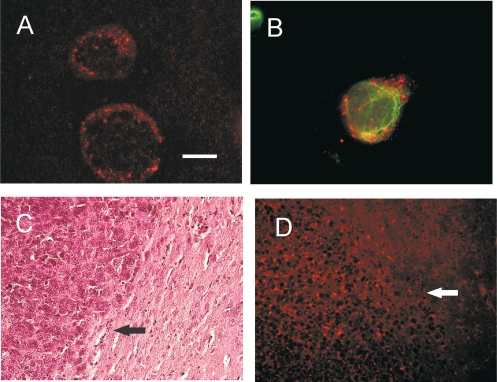
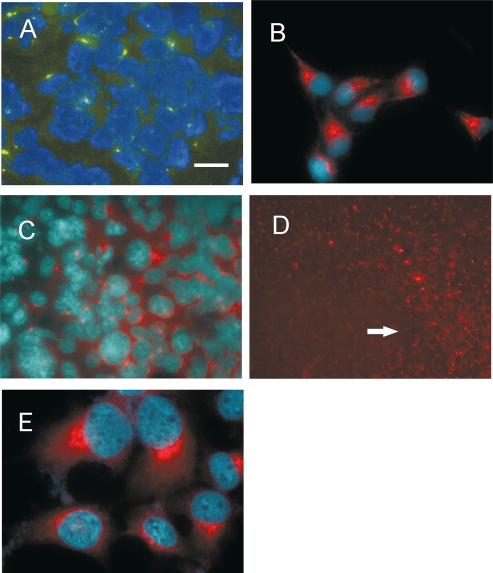
Differential diagnosis of brain tumor types is mainly based on cell morphology and could benefit from additional markers. The cAMP second-messenger system is involved in regulating cell proliferation and differentiation and is conceivably modulated during cancer transformation. The cAMP second-messenger system mainly activates protein kinases, which are in part docked to cytoskeleton, membranes, or organelles by anchoring proteins, forming protein aggregates that are detergent insoluble and not freely diffusible and that are characteristic for each cell type. The intracellular distribution of the detergent-insoluble regulatory subunits (R) of the cAMP-dependent protein kinase has been examined in mouse and rat glioma cells both in vitro and in vivo by immunohistochemistry. In normal rodent brains, the RIIalpha regulatory subunit is detergent insoluble only in ependymal cells, while in the rest of the brain it is present in soluble form. Immunohistochemistry shows that in both mouse and rat glioma cell lines, RIIalpha is mainly detergent insoluble. RIIalpha is localized close to the nucleus, associated with smooth vesicles in the trans-Golgi network area. Both paclitaxel and vinblastine cause a redistribution of RIIalpha within the cell. Under conditions that increased intracellular cAMP, apoptosis of glioma cells was observed, and it was accompanied by RIIalpha redistribution. Also in vivo, detergent-insoluble RIIalpha can be observed in mouse and rat gliomas, where it delineates the border between normal brain tissue and glioma. Therefore, intracellular distribution of detergent-insoluble RIIalpha can assist in detecting tumor cells within the brain, thus making the histologic diagnosis of brain tumors more accurate, and may represent an additional target for therapy.
Figures
Similar articles
-
Aggregates of cAMP-dependent kinase isoforms characterize different areas in the developing central nervous system of the chicken, Gallus gallus.Dev Neurosci. 2011;33(2):144-58. doi: 10.1159/000329546. Epub 2011 Aug 25. Dev Neurosci. 2011. PMID: 21876332
-
The role of protein kinase A anchoring via the RII alpha regulatory subunit in the murine immune system.J Immunol. 2005 Jun 1;174(11):6847-53. doi: 10.4049/jimmunol.174.11.6847. J Immunol. 2005. PMID: 15905526
-
Differential localization of protein kinase A type II isozymes in the Golgi-centrosomal area.Exp Cell Res. 1999 May 25;249(1):131-46. doi: 10.1006/excr.1999.4447. Exp Cell Res. 1999. PMID: 10328961
-
Patterns of cyclic AMP-dependent protein kinase gene expression during ontogeny of the murine palate.J Cell Physiol. 1995 Jun;163(3):431-40. doi: 10.1002/jcp.1041630302. J Cell Physiol. 1995. PMID: 7775586
-
Structure, function, and regulation of human cAMP-dependent protein kinases.Adv Second Messenger Phosphoprotein Res. 1997;31:191-204. doi: 10.1016/s1040-7952(97)80019-5. Adv Second Messenger Phosphoprotein Res. 1997. PMID: 9344252 Review.
Cited by
-
The Interplay of Microtubules with Mitochondria-ER Contact Sites (MERCs) in Glioblastoma.Biomolecules. 2022 Apr 12;12(4):567. doi: 10.3390/biom12040567. Biomolecules. 2022. PMID: 35454156 Free PMC article. Review.
-
Protein Kinase A Distribution in Meningioma.Cancers (Basel). 2019 Oct 29;11(11):1686. doi: 10.3390/cancers11111686. Cancers (Basel). 2019. PMID: 31671850 Free PMC article.
-
Targeting the Regulatory Subunit R2Alpha of Protein Kinase A in Human Glioblastoma through shRNA-Expressing Lentiviral Vectors.Viruses. 2021 Jul 13;13(7):1361. doi: 10.3390/v13071361. Viruses. 2021. PMID: 34372567 Free PMC article.
-
Protein kinase A regulatory subunit distribution in medulloblastoma.BMC Cancer. 2010 Apr 14;10:141. doi: 10.1186/1471-2407-10-141. BMC Cancer. 2010. PMID: 20398275 Free PMC article.
-
Protein Kinase A Distribution Differentiates Human Glioblastoma from Brain Tissue.Cancers (Basel). 2017 Dec 21;10(1):2. doi: 10.3390/cancers10010002. Cancers (Basel). 2017. PMID: 29267253 Free PMC article.
References
-
- Ohgaki H, Kleihues P. Epidemiology and etiology of gliomas. Acta Neuropathol (Berl) 2005;109:93–108. - PubMed
-
- Stork PJ, Schmitt JM. Crosstalk between cAMP and MAP kinase signalling in the regulation of cell proliferation. Trends Cell Biol. 2002;12:258–266. - PubMed
-
- Sato S, Sugimura T, Yoda K, Fujimura S. Morphological differentiation of cultured mouse glioblastoma cells induced by dibutyryl cyclic adenosine monophosphate. Cancer Res. 1975;35:2494–2499. - PubMed
-
- Moreno MJ, Ball M, Andrade MF, McDermid A, Stanimirovic DB. Insulin-like growth factor binding protein-4 (IGFBP-4) is a novel anti-angiogenic and anti-tumorigenic mediator secreted by dibutyryl cyclic AMP (dB-cAMP)-differentiated glioblastoma cells. Glia. 2006;53:845–857. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
