

Models of toxic beta-sheet channels of protegrin-1 suggest a common subunit organization motif shared with toxic alzheimer beta-amyloid ion channels
- PMID: 18708452
- PMCID: PMC2576390
- DOI: 10.1529/biophysj.108.134551
Models of toxic beta-sheet channels of protegrin-1 suggest a common subunit organization motif shared with toxic alzheimer beta-amyloid ion channels
Abstract
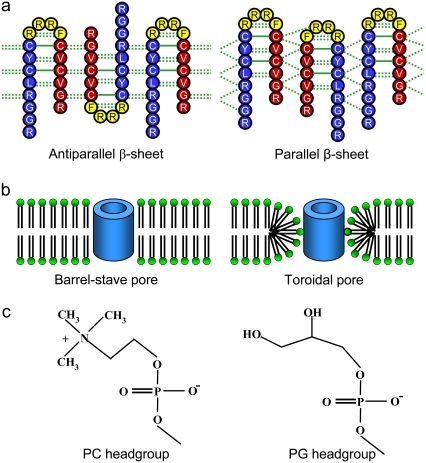
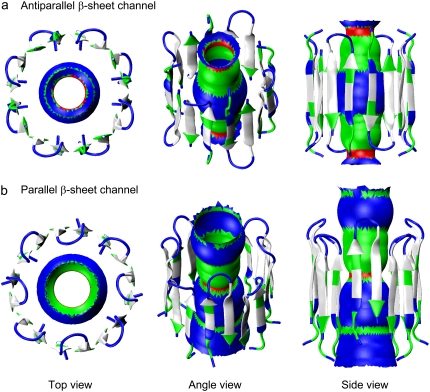
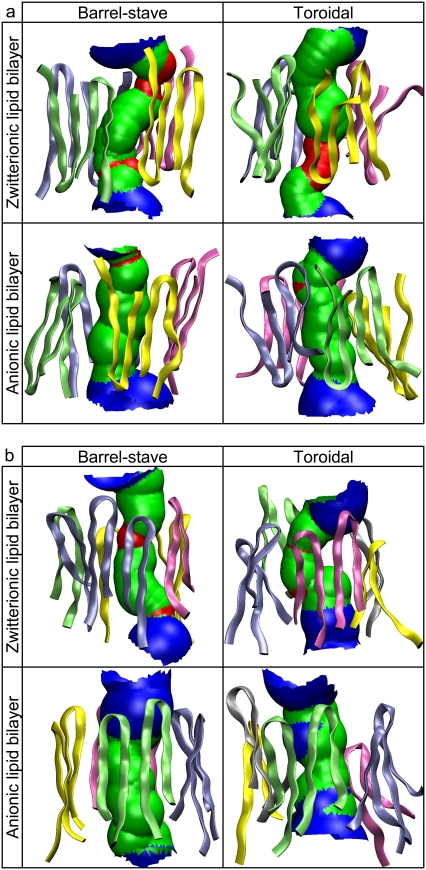
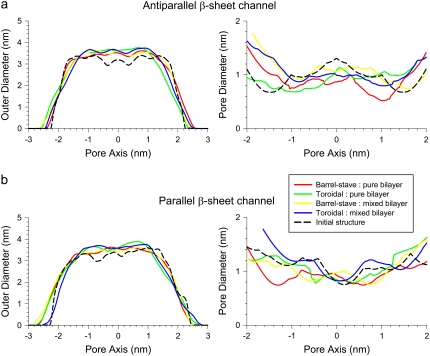
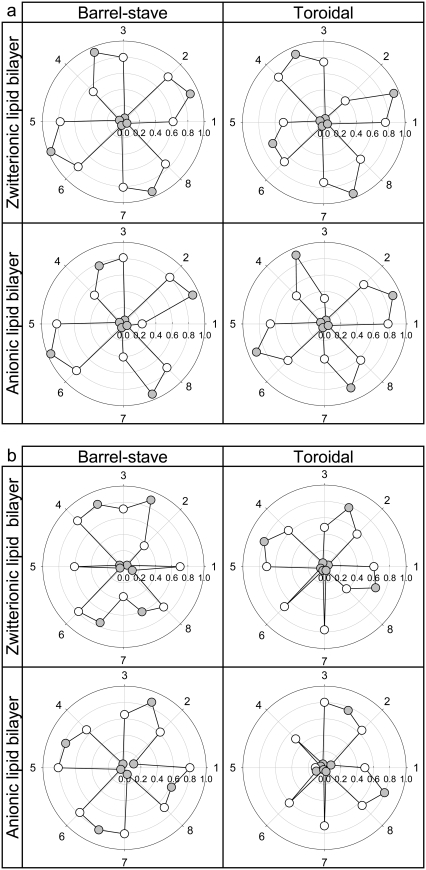
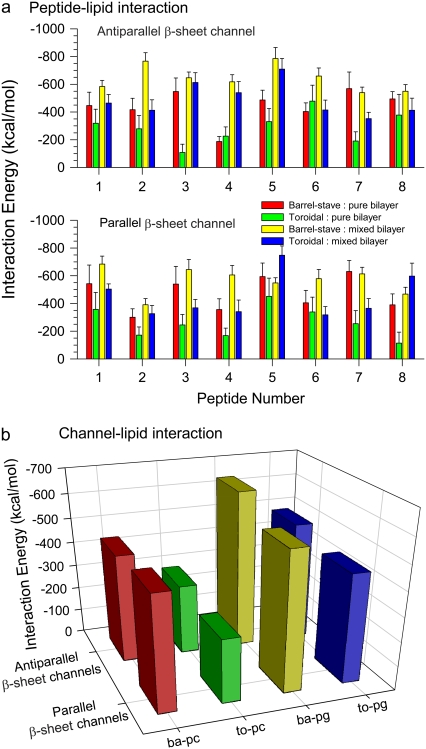
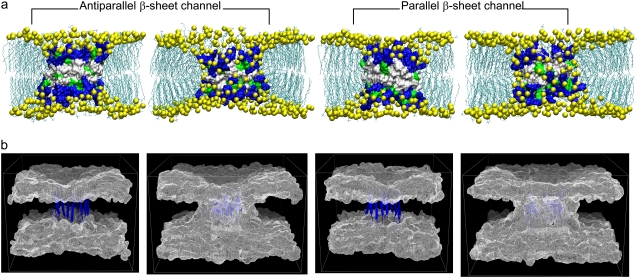
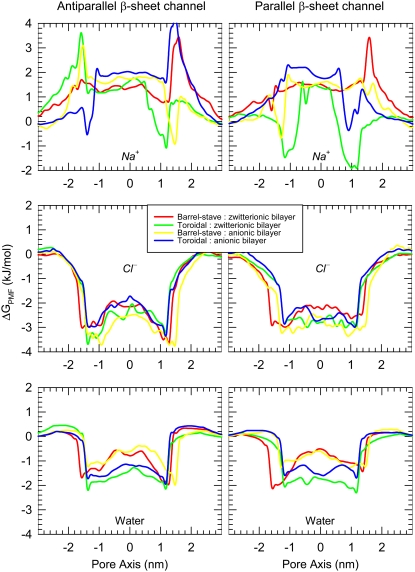
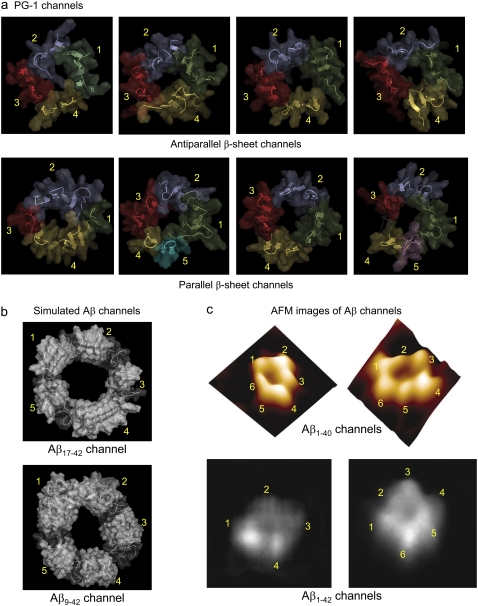
Antimicrobial peptides (AMPs) induce cytotoxicity by altering membrane permeability. The electrical properties of membrane-associated AMPs as well as their cellular effects have been extensively documented; however their three-dimensional structure is poorly understood. Gaining insight into channel structures is important to the understanding of the protegrin-1 (PG-1) and other AMP cytolytic mechanisms, and to antibiotics design. We studied the beta-sheet channels morphology using molecular dynamics simulations. We modeled PG-1 channels as intrinsic barrel-stave and toroidal membrane pores, and simulated them in zwitterionic and anionic lipid bilayers. PG-1 channels consist of eight beta-hairpins in a consecutive NCCN (N and C represent the beta-hairpin's N- and C-termini) packing organization yielding antiparallel and parallel beta-sheet channels. Both channels preserve the toroidal, but not the barrel-stave pores. The two lipid leaflets of the bilayer bend toward each other at the channels' edges, producing a semitoroidal pore with the outward-pointing hydrophobic residues preventing the polar lipid headgroups from moving to the bilayer center. In all simulated lipid environments, PG-1 channels divide into four or five beta-sheet subunits consisting of single or dimeric beta-hairpins. The channel morphology with subunit organization is consistent with the four to five subunits observed by NMR in the POPE/POPG bilayer. Remarkably, a beta-sheet subunit channel motif is in agreement with Alzheimer ion channels modeled using the universal U-shape beta-strand-turn-beta-strand structure, as well as with high resolution atomic force microscopy images of beta-amyloid channels with four to six subunits. Consistent with the toxic beta-amyloid channels that are ion-conducting, the PG-1 channels permeate anions.
Figures
References
-
- Sitaram, N., and R. Nagaraj. 2002. Host-defense antimicrobial peptides: importance of structure for activity. Curr. Pharm. Des. 8:727–742. - PubMed
-
- Hancock, R. E., and A. Rozek. 2002. Role of membranes in the activities of antimicrobial cationic peptides. FEMS Microbiol. Lett. 206:143–149. - PubMed
-
- Kourie, J. I., and A. A. Shorthouse. 2000. Properties of cytotoxic peptide-formed ion channels. Am. J. Physiol. Cell Physiol. 278:C1063–C1087. - PubMed
-
- Zasloff, M. 2002. Antimicrobial peptides of multicellular organisms. Nature. 415:389–395. - PubMed
-
- Sokolov, Y., T. Mirzabekov, D. W. Martin, R. I. Lehrer, and B. L. Kagan. 1999. Membrane channel formation by antimicrobial protegrins. Biochim. Biophys. Acta. 1420:23–29. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
