

Neither helix in the coiled coil region of the axle of F1-ATPase plays a significant role in torque production
- PMID: 18708468
- PMCID: PMC2576389
- DOI: 10.1529/biophysj.108.140061
Neither helix in the coiled coil region of the axle of F1-ATPase plays a significant role in torque production
Abstract
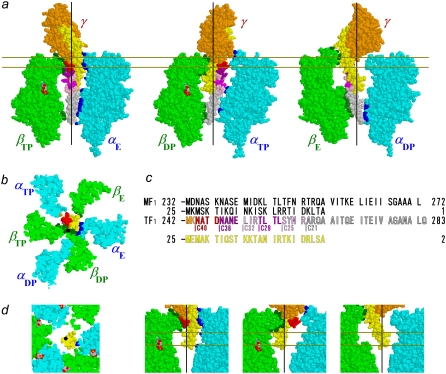
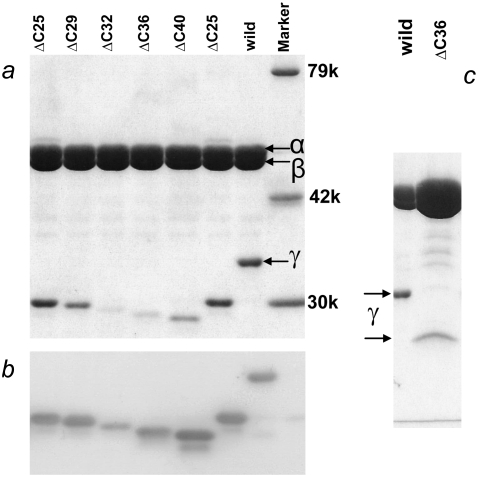
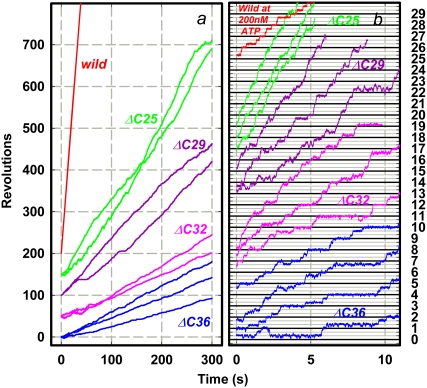
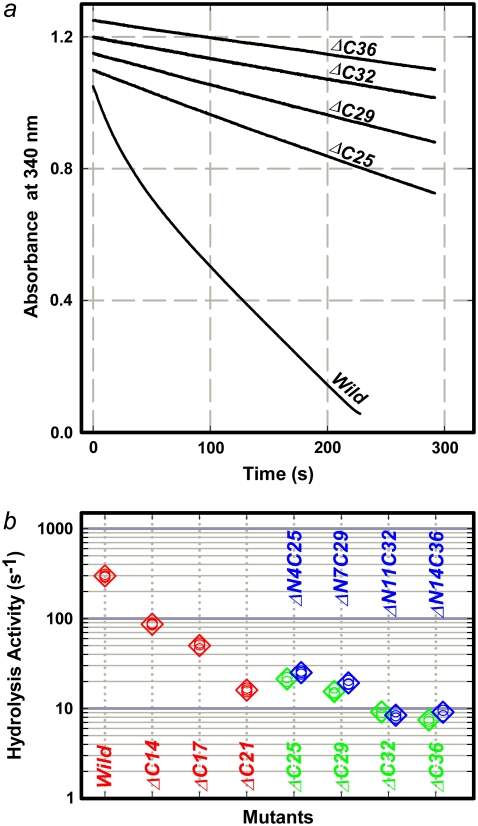
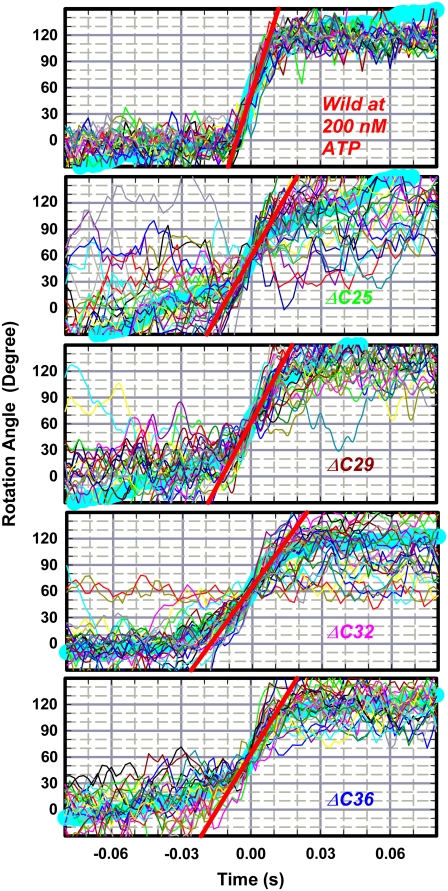
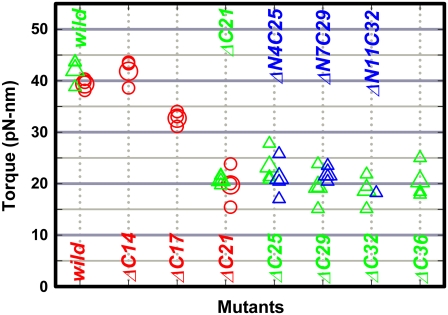
F(1)-ATPase is an ATP-driven rotary molecular motor in which the central gamma-subunit rotates inside the cylinder made of alpha(3)beta(3) subunits. The amino and carboxy termini of the gamma-subunit form the axle, an alpha-helical coiled coil that deeply penetrates the stator cylinder. We previously truncated the axle step by step, starting with the longer carboxy terminus and then cutting both termini at the same levels, resulting in a slower yet considerably powerful rotation. Here we examine the role of each helix by truncating only the carboxy terminus by 25-40 amino-acid residues. Longer truncation impaired the stability of the motor complex severely: 40 deletions failed to yield rotating the complex. Up to 36 deletions, however, the mutants produced an apparent torque at nearly half of the wild-type torque, independent of truncation length. Time-averaged rotary speeds were low because of load-dependent stumbling at 120 degrees intervals, even with saturating ATP. Comparison with our previous work indicates that half the normal torque is produced at the orifice of the stator. The very tip of the carboxy terminus adds the other half, whereas neither helix in the middle of the axle contributes much to torque generation and the rapid progress of catalysis. None of the residues of the entire axle played a specific decisive role in rotation.
Figures
References
-
- Kagawa, Y., and E. Racker. 1966. Partial resolution of the enzymes catalyzing oxidative phosphorylation. IX. Reconstruction of oligomycin-sensitive adenosine triphosphatase. J. Biol. Chem. 241:2467–2474. - PubMed
-
- Catteral, W. A., and P. L. Pedersen. 1971. Adenosine triphosphatase from rat liver mitochondria. I. Purification, homogeneity, and physical properties. J. Biol. Chem. 246:4987–4994. - PubMed
-
- Yoshida, M., E. Muneyuki, and T. Hisabori. 2001. ATP synthase—a marvelous rotary engine of the cell. Nat. Rev. Mol. Cell Biol. 2:669–677. - PubMed
-
- Boyer, P. D., and W. E. Kohlbrenner. 1981. The present status of the binding-change mechanism and its relation to ATP formation by chloroplasts. In Energy Coupling in Photosynthesis. B. R. Selman and S. Selman-Reimer, editors. Elsevier, Amsterdam, The Netherlands. 231–240.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
