

doi: 10.1128/JB.00934-08.
Epub 2008 Aug 15.
Interaction of penicillin-binding protein 2 with soluble lytic transglycosylase B1 in Pseudomonas aeruginosa
Affiliations
- PMID: 18708507
- PMCID: PMC2566182
- DOI: 10.1128/JB.00934-08
Item in Clipboard
Interaction of penicillin-binding protein 2 with soluble lytic transglycosylase B1 in Pseudomonas aeruginosa
J Bacteriol.
2008 Oct.
Abstract
Soluble lytic transglycosylase B1 from Pseudomonas aeruginosa was coupled to Sepharose and used to immobilize interaction partners from membrane protein extracts. Penicillin-binding protein 2 (PBP2) was identified as a binding partner, suggesting that the two proteins function together in the biosynthesis of peptidoglycan. By use of an engineered truncated derivative, the N-terminal module of PBP2 was found to confer the binding properties.
Figures
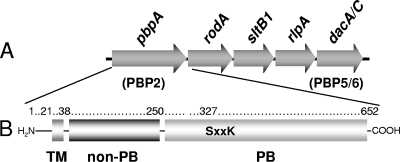
Organization of the gene cluster harboring pbpA and modular structure of PBP2 from P. aeruginosa. (A) The cluster of genes at the pbpA (PBP2) locus on the P. aeruginosa chromosome includes dacA-dacC (dacA/C) and sltB1, encoding PBP5-PBP6 and SltB1, respectively. (B) The structure of PBP2 is comprised of an N-terminal transmembrane anchor (TM; residues 1 to 38), a non-PB module (residues 39 to 250), and a C-terminal PB module (residues 256 to 652) connected by a short linker sequence. Depicted within the PB module is the Ser-X-X-Lys consensus sequence involving Ser327 as the catalytic residue and site of penicillin binding.
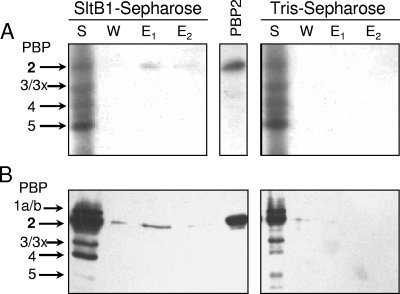
PBP assays of fractions from affinity chromatography of membrane proteins on SltB-Sepharose. A P. aeruginosa PA01 membrane-protein extract (S) was applied to either SltB1-Sepharose or a control column of Tris-Sepharose in 10 mM Tris-HCl, 10 mM MgCl2, 50 mM NaCl, and 0.05% Triton X-100. After the flowthrough fraction was collected, the resins were washed with the same buffer containing 150 mM NaCl (W) and then eluted with buffer containing 1 M NaCl (E1) and 5 mM EGTA (E2). Samples (30 μl) of the fractions were incubated with [3H]penicillin G (A) or biotinylated ampicillin (B) and analyzed by SDS-PAGE as described in Materials and Methods. The PBPs were visualized by autoradiography (A) or chemiluminescence (B). A sample of purified P. aeruginosa PBP2 (PBP2) was applied as a positive control, and the assignment of the other PBPs was based on apparent molecular masses.
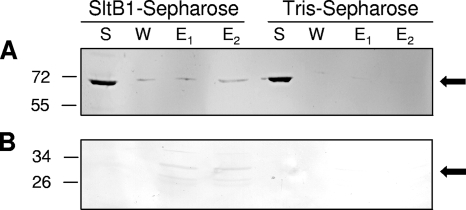
Immunoblots of fractions from affinity chromatography of PBP2 applied to SltB1-Sepharose and Tris-Sepharose. Purified preparations of recombinant PBP2 (A) and the isolated non-PB module (B) were applied to either SltB1-Sepharose or a control Tris-Sepharose column. Conditions of the affinity chromatography and SDS-PAGE were as described in the legend to Fig. 2. Following electrophoresis, the recombinant proteins were detected by Western immunoblot analysis using an anti-six-His antibody as described in Materials and Methods. The positions of molecular mass markers (in kilodaltons) are indicated on the left, and the arrows denote the positions of PBP2 (top) and the non-PB module (bottom).
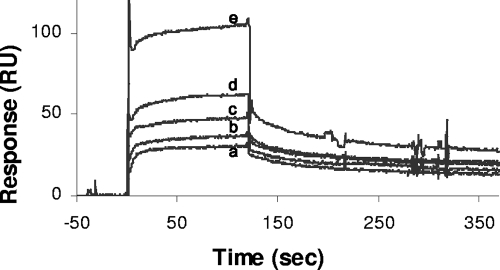
SPR analysis of interaction between P. aeruginosa SltB1 and PBP2. The soluble derivative of PBP2, sPBP2, in 10 mM HEPES, 150 mM NaCl, and 0.005% Tween 20 (pH 7.4) was applied to SltB1 immobilized on a CM5 sensor chip at a flow rate of 100 μl/min. The representative sensorgrams show the responses to injections of sPBP2 at concentrations of 230 nM (a), 460 nM (b), 920 nM (c), 1,840 nM (d), and 3,690 nM (e) using a Biacore 2000 spectrometer.
Similar articles
-
Calcium-dependent complex formation between PBP2 and lytic transglycosylase SltB1 of Pseudomonas aeruginosa.Microb Drug Resist. 2012 Jun;18(3):298-305. doi: 10.1089/mdr.2012.0006. Epub 2012 Mar 20. Microb Drug Resist. 2012. PMID: 22432706
-
Substrate binding affinity of Pseudomonas aeruginosa membrane-bound lytic transglycosylase B by hydrogen-deuterium exchange MALDI MS.Biochemistry. 2004 Sep 7;43(35):11275-82. doi: 10.1021/bi049496d. Biochemistry. 2004. PMID: 15366937
-
Lytic transglycosylase Slt of Pseudomonas aeruginosa as a periplasmic hub protein.Protein Sci. 2024 Jul;33(7):e5038. doi: 10.1002/pro.5038. Protein Sci. 2024. PMID: 38864725 Free PMC article.
-
The penicillin-binding proteins: structure and role in peptidoglycan biosynthesis.FEMS Microbiol Rev. 2008 Mar;32(2):234-58. doi: 10.1111/j.1574-6976.2008.00105.x. Epub 2008 Feb 11. FEMS Microbiol Rev. 2008. PMID: 18266856 Review.
-
Lytic transglycosylases.EXS. 1996;75:425-9. doi: 10.1007/978-3-0348-9225-4_21. EXS. 1996. PMID: 8765311 Review.
Cited by
-
Changes to its peptidoglycan-remodeling enzyme repertoire modulate β-lactam resistance in Pseudomonas aeruginosa.Antimicrob Agents Chemother. 2013 Jul;57(7):3078-84. doi: 10.1128/AAC.00268-13. Epub 2013 Apr 22. Antimicrob Agents Chemother. 2013. PMID: 23612194 Free PMC article.
-
Fluorescence Assessment of the AmpR-Signaling Network of Pseudomonas aeruginosa to Exposure to β-Lactam Antibiotics.ACS Chem Biol. 2020 May 15;15(5):1184-1194. doi: 10.1021/acschembio.9b00875. Epub 2020 Feb 10. ACS Chem Biol. 2020. PMID: 31990176 Free PMC article.
-
Lytic transglycosylases: concinnity in concision of the bacterial cell wall.Crit Rev Biochem Mol Biol. 2017 Oct;52(5):503-542. doi: 10.1080/10409238.2017.1337705. Epub 2017 Jun 23. Crit Rev Biochem Mol Biol. 2017. PMID: 28644060 Free PMC article. Review.
-
The Legionella pneumophila EnhC protein interferes with immunostimulatory muramyl peptide production to evade innate immunity.Cell Host Microbe. 2012 Aug 16;12(2):166-76. doi: 10.1016/j.chom.2012.06.004. Cell Host Microbe. 2012. PMID: 22901537 Free PMC article.
-
Penicillin-binding protein 5 can form a homo-oligomeric complex in the inner membrane of Escherichia coli.Protein Sci. 2011 Sep;20(9):1520-9. doi: 10.1002/pro.677. Epub 2011 Jul 13. Protein Sci. 2011. PMID: 21674665 Free PMC article.
References
-
- Alaedini, A., and R. A. Day. 1999. Identification of two penicillin-binding multienzyme complexes in Haemophilus influenzae. Biochem. Biophys. Res. Commun. 264191-195. - PubMed
-
- Bayer, E. A., J. P. Belaich, Y. Shoham, and R. Lamed. 2004. The cellulosomes: multienzyme machines for degradation of plant cell wall polysaccharides. Annu. Rev. Microbiol. 58521-554. - PubMed
-
- Bertsche, U., T. Kast, B. Wolf, C. Fraipont, M. E. G. Aarsman, K. Kannenberg, M. von Reschenberg, M. Nguyen-Distèche, T. den Blaauwen, J.-V. Höltje, and W. Vollmer. 2006. Interaction between two murein (peptidoglycan) synthases, PBP 3 and PBP 1B, in Escherichia coli. Mol. Microbiol. 61675-690. - PubMed
-
- Bhardwaj, S., and R. A. Day. 1997. Detection of intra-cellular protein-protein interactions: penicillin interactive proteins and morphogene proteins, p. 469-480. In D. R. Marshak (ed.), Techniques in protein chemistry, vol. 8. Academic Press, San Diego, CA.
-
- Blackburn, N. T., and A. J. Clarke. 2002. Characterization of soluble and membrane-bound family 3 lytic transglycosylases from Pseudomonas aeruginosa. Biochemistry 411001-1013. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases