

Replication termination mechanism as revealed by Tus-mediated polar arrest of a sliding helicase
- PMID: 18708526
- PMCID: PMC2529109
- DOI: 10.1073/pnas.0805898105
Replication termination mechanism as revealed by Tus-mediated polar arrest of a sliding helicase
Abstract
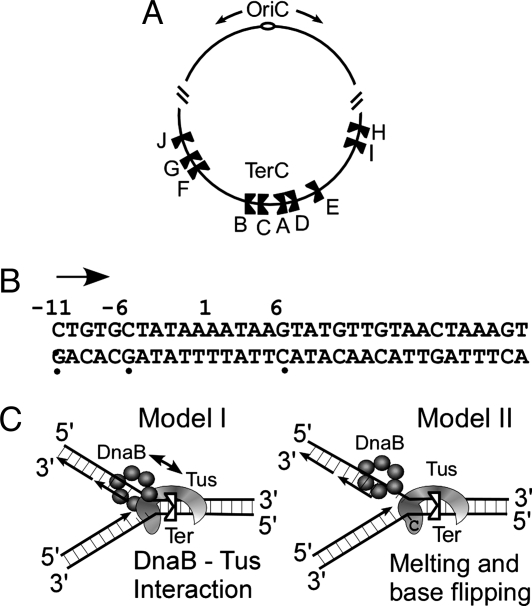
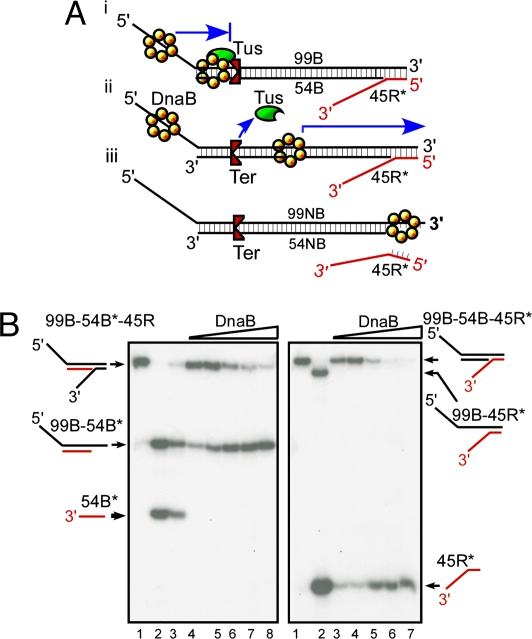
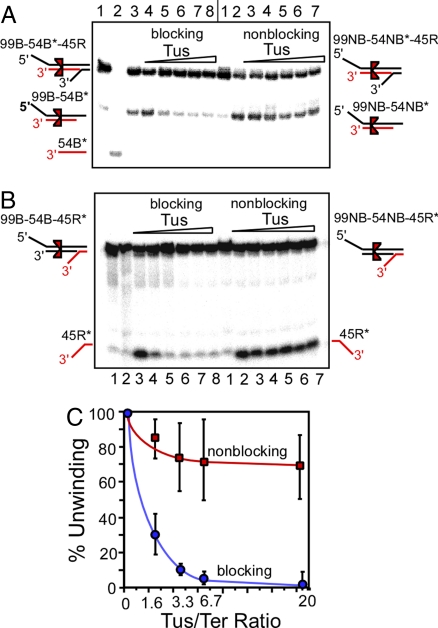
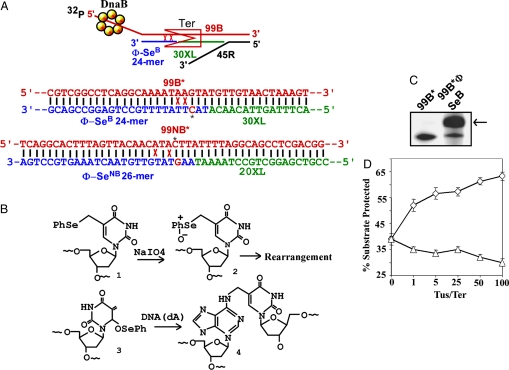
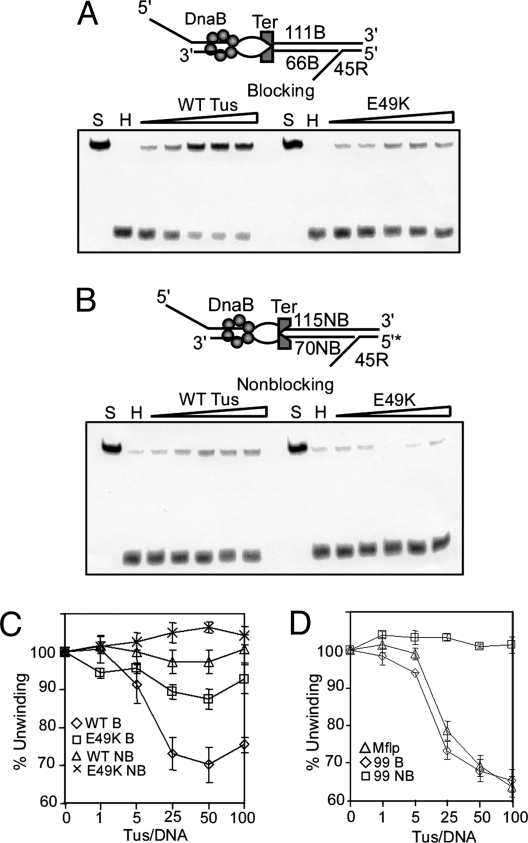
The replication terminator protein Tus of Escherichia coli promotes polar fork arrest at sequence-specific replication termini (Ter) by antagonizing DNA unwinding by the replicative helicase DnaB. Here, we report that Tus is also a polar antitranslocase. We have used this activity as a tool to uncouple helicase arrest at a Tus-Ter complex from DNA unwinding and have shown that helicase arrest occurred without the generation of a DNA fork or a bubble of unpaired bases at the Tus-Ter complex. A mutant form of Tus, which reduces DnaB-Tus interaction but not the binding affinity of Tus for Ter DNA, was also defective in arresting a sliding DnaB. A model of polar fork arrest that proposes melting of the Tus-Ter complex and flipping of a conserved C residue of Ter at the blocking but not the nonblocking face has been reported. The model suggests that enhanced stability of Tus-Ter interaction caused by DNA melting and capture of a flipped base by Tus generates polarity strictly by enhanced protein-DNA interaction. In contrast, the observations presented here show that polarity of helicase and fork arrest in vitro is generated by a mechanism that not only involves interaction between the terminator protein and the arrested enzyme but also of Tus with Ter DNA, without any melting and base flipping in the termination complex.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Bastia D, Mohanty BK. In: DNA Replication in Eukaryotic Cells. DePamphilis M, editor. Plainview, NY: Cold Spring Harbor Lab Press; 1996. pp. 177–215.
-
- Bastia D, Mohanty B. K. In: DNA Replication and Human Disease. DePamphilis M, editor. Plainview, NY: Cold Spring Harbor Lab Press; 2006. pp. 155–174.
-
- Rothstein R, Michel B, Gangloff S. Replication fork pausing and recombination or “gimme a break”. Genes Dev. 2000;14:1–10. - PubMed
-
- Brewer BJ, Fangman WL. A replication fork barrier at the 3′ end of yeast ribosomal RNA genes. Cell. 1988;55:637–643. - PubMed
-
- Dalgaard JZ, Klar AJ. Orientation of DNA replication establishes mating-type switching pattern in S. pombe. Nature. 1999;400:181–184. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
