

Review
doi: 10.1101/gad.1693208.
Plant hormone receptors: new perceptions
Affiliations
- PMID: 18708574
- PMCID: PMC2735353
- DOI: 10.1101/gad.1693208
Item in Clipboard
Review
Plant hormone receptors: new perceptions
Genes Dev.
.
Abstract
Plant growth and development require the integration of a variety of environmental and endogenous signals that, together with the intrinsic genetic program, determine plant form. Central to this process are several growth regulators known as plant hormones or phytohormones. Despite decades of study, only recently have receptors for several of these hormones been identified, revealing novel mechanisms for perceiving chemical signals and providing plant biologists with a much clearer picture of hormonal control of growth and development.
Figures
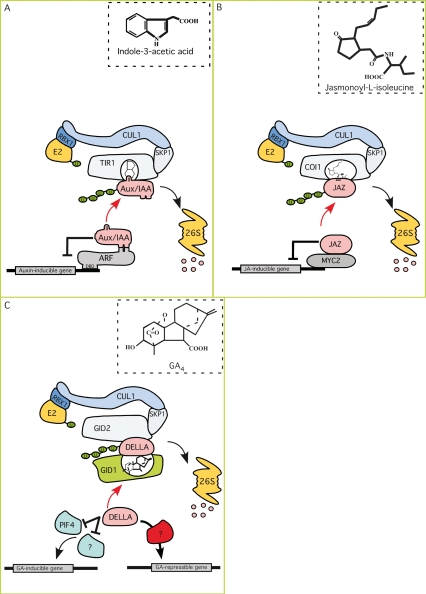
Ubiquitin-ligase based receptors. (A) Under subthreshold auxin concentrations, auxin-responsive genes are repressed by Aux/IAA proteins heterodimerizing with ARF transcription factors. Upon an auxin stimulus, the TIR1 F-box protein subunit of the SCFTIR1 ubiquitin-ligase binds auxin, enabling the recruitment of Aux/IAA proteins to the SCF complex for ubiquitination. Subsequent Aux/IAA degradation by the 26S proteasome derepresses the ARF transcription factors. (B) JAZ proteins negatively regulate jasmonate response by repressing MYC2 transcriptional activity. Upon binding jasmonate, the SCFCOI1 ubiquitin-ligase targets JAZ proteins for ubiquitin-mediated proteolysis, derepressing MYC2. (C) DELLA proteins repress GA response by negatively regulating PIF3, PIF4, and presumably other transcription factors that control the expression of GA-inducible genes. DELLA proteins also promote the expression of several GA-repressible genes, several of which encode GA biosynthetic enzymes and components of the response pathway including the GID1 receptors. Upon binding GA, the GID1 receptor interacts with DELLA. The GID1-GA-DELLA complex is then recognized by the SCFGID2 ubiquitin-ligase, which targets DELLA for ubiquitin-mediated degradation. The red arrows in A–C indicate the effects of the respective hormone on the Aux/IAA, JAZ, and DELLA repressors.
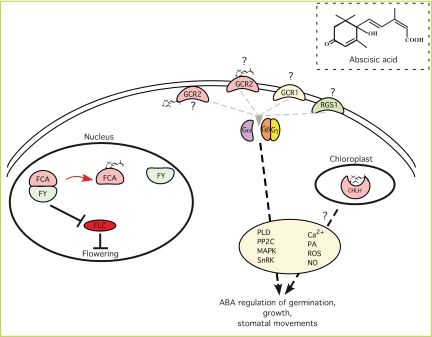
ABA receptors act at different cellular locations. The RNA-binding protein FCA acts in the nucleus together with the mRNA 3′-end processing factor FY to control the abundance of FLC, an inhibitor of flowering. Upon binding ABA, FCA and FY dissociate, leading to the accumulation of FLC and inhibition of flowering. In the chloroplast, the Mg-chelatase H subunit (CHLH) binds to ABA and acts as a positive regulator of ABA signaling by an unknown mechanism. This function appears to be separable from CHLH's role in plastid-to-nucleus signaling and chlorophyll synthesis. GPCRs have been proposed to function as ABA receptors on the plasma membrane. Genetic studies have implicated GCR1 and RGS1 as potential ABA receptors, but ABA binding activity has not been demonstrated. The GPCR-related protein GCR2 does bind ABA. However, it is currently unclear whether or not GCR2 is a canonical serpentine GPCR or a peripheral membrane protein. Genetic studies have implicated G-proteins and several potential downstream effectors and secondary messengers in ABA response. (PLD) Phospholipase D; (PP2C) protein phosphatase 2C; (MAPK) mitogen-activated kinase; (SnRK) SNF1-related kinase; (PA) phosphatidic acid; (ROS) reactive oxygen species; (NO) nitric oxide.
Similar articles
-
Plant hormone receptors: perception is everything.Genes Dev. 2006 Aug 1;20(15):1998-2008. doi: 10.1101/gad.1432806. Genes Dev. 2006. PMID: 16882977 Review.
-
Signs of change: hormone receptors that regulate plant development.Development. 2006 May;133(10):1857-69. doi: 10.1242/dev.02359. Development. 2006. PMID: 16651539 Review.
-
Recent advances in the study of mechanisms of action of phytohormones.Biochemistry (Mosc). 2004 Mar;69(3):233-47. doi: 10.1023/b:biry.0000022053.73461.cd. Biochemistry (Mosc). 2004. PMID: 15061689 Review.
-
Ligand-receptor interactions in plant hormone signaling.Plant J. 2021 Jan;105(2):290-306. doi: 10.1111/tpj.15115. Epub 2021 Jan 8. Plant J. 2021. PMID: 33278046 Review.
-
Receptors and signalling components of plant hormones.J Recept Signal Transduct Res. 1999 Jan-Jul;19(1-4):41-58. doi: 10.3109/10799899909036636. J Recept Signal Transduct Res. 1999. PMID: 10071749 Review.
Cited by
-
Structural insights into AtABCG25, an angiosperm-specific abscisic acid exporter.Plant Commun. 2024 Jan 8;5(1):100776. doi: 10.1016/j.xplc.2023.100776. Epub 2023 Dec 3. Plant Commun. 2024. PMID: 38050355 Free PMC article.
-
Isolation and characterization of gene encoding G protein α subunit protein responsive to plant hormones and abiotic stresses in Brassica napus.Mol Biol Rep. 2010 Dec;37(8):3957-65. doi: 10.1007/s11033-010-0054-x. Epub 2010 Mar 18. Mol Biol Rep. 2010. PMID: 20238175
-
Gene expression analysis and halting of ethylene receptors signaling pinpoint ethylene as a positive regulator of direct somatic embryogenesis in Coffea arabica.Planta. 2025 Aug 8;262(3):74. doi: 10.1007/s00425-025-04798-8. Planta. 2025. PMID: 40779278
-
Jasmonate signalling in Arabidopsis involves SGT1b-HSP70-HSP90 chaperone complexes.Nat Plants. 2015;1:15049. doi: 10.1038/nplants.2015.49. Epub 2015 Apr 27. Nat Plants. 2015. PMID: 27054042 Free PMC article.
-
A maize phytochrome-interacting factor 3 improves drought and salt stress tolerance in rice.Plant Mol Biol. 2015 Mar;87(4-5):413-28. doi: 10.1007/s11103-015-0288-z. Epub 2015 Jan 31. Plant Mol Biol. 2015. PMID: 25636202
References
-
- Benavente L.M., Alonso J.M. Molecular mechanisms of ethylene signaling in Arabidopsis. Mol. Biosyst. 2006;2:165–173. - PubMed
-
- Chini A., Fonseca S., Fernandez G., Adie B., Chico J.M., Lorenzo O., Garcia-Casado G., Lopez-Vidriero I., Lozano F.M., Ponce M.R., et al. The JAZ family of repressors is the missing link in jasmonate signalling. Nature. 2007;448:666–671. - PubMed
-
- Chow B., McCourt P. Plant hormone receptors: Perception is everything. Genes & Dev. 2006;20:1998–2008. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources