

NFAT regulates induction of COX-2 and apoptosis of keratinocytes in response to ultraviolet radiation exposure
- PMID: 18708588
- PMCID: PMC2671982
- DOI: 10.1096/fj.08-113076
NFAT regulates induction of COX-2 and apoptosis of keratinocytes in response to ultraviolet radiation exposure
Abstract
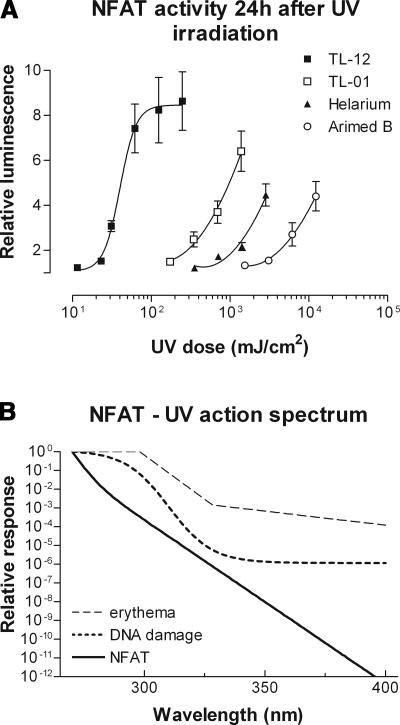
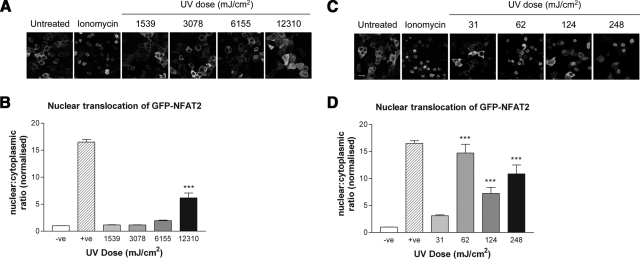
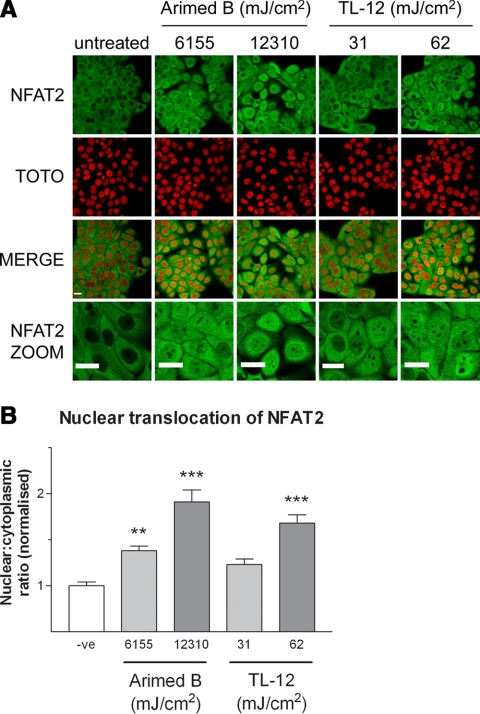
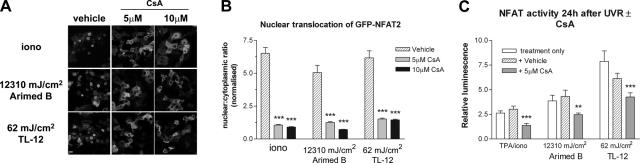
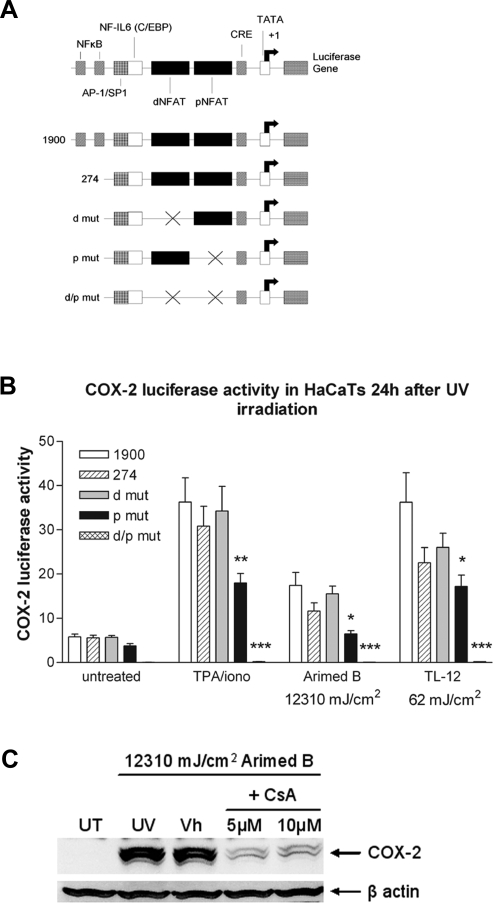
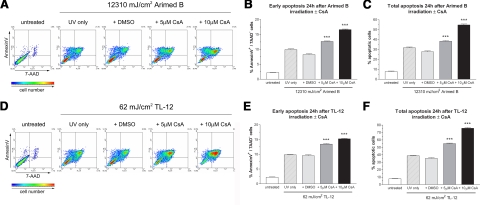
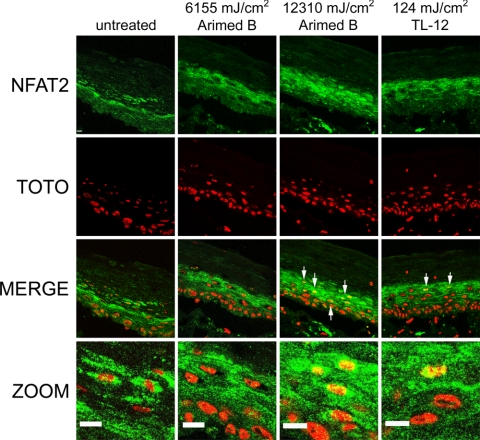
The nuclear factor of activated T cells (NFAT) transcription factors are regulated by calcium/calcineurin signals and play important roles in T cells, muscle, bone, and neural tissue. NFAT is expressed in the epidermis, and although recent data suggest that NFAT is involved in the skin's responses to ultraviolet radiation (UVR), the wavelengths of radiation that activate NFAT and the biological function of UV-activated NFAT remain undefined. We demonstrate that NFAT transcriptional activity is preferentially induced by UVB wavelengths in keratinocytes. The derived action spectrum for NFAT activation indicates that NFAT transcriptional activity is inversely associated with wavelength. UVR also evoked NFAT2 nuclear translocation in a parallel wavelength-dependent fashion and both transcriptional activation and nuclear translocation were inhibited by the calcineurin inhibitor cyclosporin A. UVR also evoked NFAT2 nuclear translocation in three-dimensional skin equivalents. Evidence suggests that COX-2 contributes to UV-induced carcinogenesis. Inhibiting UV-induced NFAT activation in keratinocytes, reduced COX-2 protein induction, and increased UV-induced apoptosis. COX-2 luciferase reporters lacking functional NFAT binding sites were less responsive to UVR, highlighting that NFAT is required for UV-induced COX-2 induction. Taken together, these data suggest that the proinflammatory, antiapoptotic, and procarcinogenic functions of UV-activated COX-2 may be mediated, in part, by upstream NFAT signaling.
Figures
References
-
- Shaw J P, Utz P J, Durand D B, Toole J J, Emmel E A, Crabtree G R. Identification of a putative regulator of early T cell activation genes. Science. 1988;241:202–205. - PubMed
-
- Kiani A, Rao A, Aramburu J. Manipulating immune responses with immunosuppressive agents that target NFAT. Immunity. 2000;12:359–372. - PubMed
-
- Hill-Eubanks D C, Gomez M F, Stevenson A S, Nelson M T. NFAT regulation in smooth muscle. Trends Cardiovasc Med. 2003;13:56–62. - PubMed
-
- Schulz R A, Yutzey K E. Calcineurin signaling and NFAT activation in cardiovascular and skeletal muscle development. Dev Biol. 2004;266:1–16. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
