

Citrullination of CXCL8 by peptidylarginine deiminase alters receptor usage, prevents proteolysis, and dampens tissue inflammation
- PMID: 18710930
- PMCID: PMC2526203
- DOI: 10.1084/jem.20080305
Citrullination of CXCL8 by peptidylarginine deiminase alters receptor usage, prevents proteolysis, and dampens tissue inflammation
Abstract
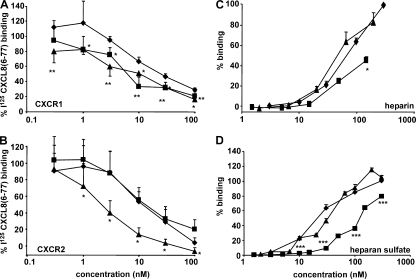
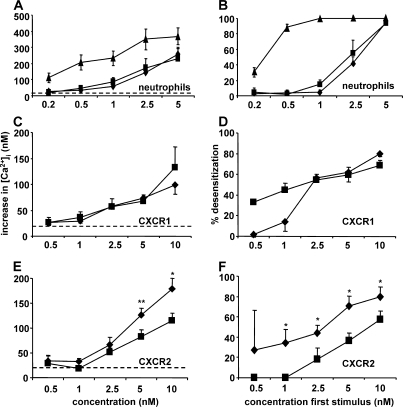
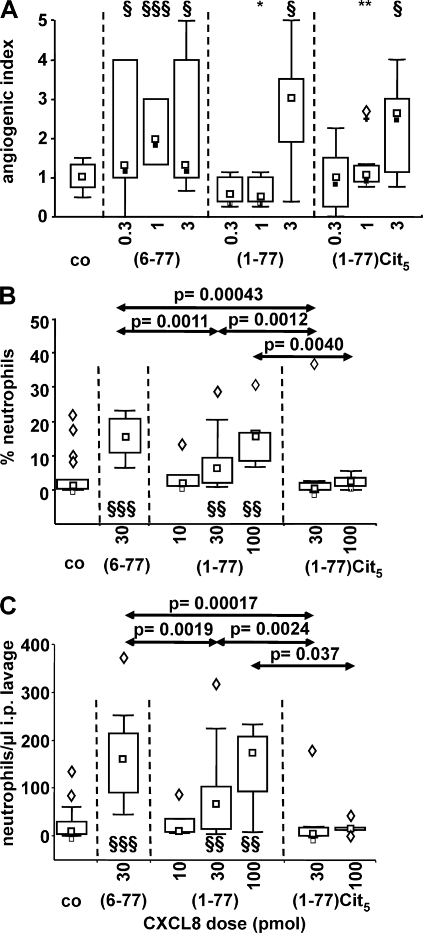
Biological functions of proteins are influenced by posttranslational modifications such as on/off switching by phosphorylation and modulation by glycosylation. Proteolytic processing regulates cytokine and chemokine activities. In this study, we report that natural posttranslational citrullination or deimination alters the biological activities of the neutrophil chemoattractant and angiogenic cytokine CXCL8/interleukin-8 (IL-8). Citrullination of arginine in position 5 was discovered on 14% of natural leukocyte-derived CXCL8(1-77), generating CXCL8(1-77)Cit(5). Peptidylarginine deiminase (PAD) is known to citrullinate structural proteins, and it may initiate autoimmune diseases. PAD efficiently and site-specifically citrullinated CXCL5, CXCL8, CCL17, CCL26, but not IL-1beta. In comparison with CXCL8(1-77), CXCL8(1-77)Cit(5) had reduced affinity for glycosaminoglycans and induced less CXCR2-dependent calcium signaling and extracellular signal-regulated kinase 1/2 phosphorylation. In contrast to CXCL8(1-77), CXCL8(1-77)Cit(5) was resistant to thrombin- or plasmin-dependent potentiation into CXCL8(6-77). Upon intraperitoneal injection, CXCL8(6-77) was a more potent inducer of neutrophil extravasation compared with CXCL8(1-77). Despite its retained chemotactic activity in vitro, CXCL8(1-77)Cit(5) was unable to attract neutrophils to the peritoneum. Finally, in the rabbit cornea angiogenesis assay, the equally potent CXCL8(1-77) and CXCL8(1-77)Cit(5) were less efficient angiogenic molecules than CXCL8(6-77). This study shows that PAD citrullinates the chemokine CXCL8, and thus may dampen neutrophil extravasation during acute or chronic inflammation.
Figures




Similar articles
-
Posttranslational modification of the NH2-terminal region of CXCL5 by proteases or peptidylarginine Deiminases (PAD) differently affects its biological activity.J Biol Chem. 2010 Sep 24;285(39):29750-9. doi: 10.1074/jbc.M110.119388. Epub 2010 Jul 14. J Biol Chem. 2010. PMID: 20630876 Free PMC article.
-
Citrullination and proteolytic processing of chemokines by Porphyromonas gingivalis.Infect Immun. 2014 Jun;82(6):2511-9. doi: 10.1128/IAI.01624-14. Epub 2014 Mar 31. Infect Immun. 2014. PMID: 24686061 Free PMC article.
-
Citrullination of CXCL10 and CXCL11 by peptidylarginine deiminase: a naturally occurring posttranslational modification of chemokines and new dimension of immunoregulation.Blood. 2008 Oct 1;112(7):2648-56. doi: 10.1182/blood-2008-04-149039. Epub 2008 Jul 21. Blood. 2008. PMID: 18645041
-
Peptidylarginine deiminases in citrullination, gene regulation, health and pathogenesis.Biochim Biophys Acta. 2013 Oct;1829(10):1126-35. doi: 10.1016/j.bbagrm.2013.07.003. Epub 2013 Jul 13. Biochim Biophys Acta. 2013. PMID: 23860259 Free PMC article. Review.
-
Citrullination and PAD Enzyme Biology in Type 1 Diabetes - Regulators of Inflammation, Autoimmunity, and Pathology.Front Immunol. 2021 Jun 1;12:678953. doi: 10.3389/fimmu.2021.678953. eCollection 2021. Front Immunol. 2021. PMID: 34140951 Free PMC article. Review.
Cited by
-
Citrullination of proteins: a common post-translational modification pathway induced by different nanoparticles in vitro and in vivo.Nanomedicine (Lond). 2012 Aug;7(8):1181-95. doi: 10.2217/nnm.11.177. Epub 2012 May 25. Nanomedicine (Lond). 2012. PMID: 22625207 Free PMC article.
-
Citrullinated Autoantigens: From Diagnostic Markers to Pathogenetic Mechanisms.Clin Rev Allergy Immunol. 2015 Oct;49(2):232-9. doi: 10.1007/s12016-014-8459-2. Clin Rev Allergy Immunol. 2015. PMID: 25355199 Review.
-
Rheumatoid arthritis and citrullination.Curr Opin Rheumatol. 2018 Jan;30(1):72-78. doi: 10.1097/BOR.0000000000000452. Curr Opin Rheumatol. 2018. PMID: 28937414 Free PMC article. Review.
-
CXCL8 and its cognate receptors in melanoma progression and metastasis.Future Oncol. 2010 Jan;6(1):111-6. doi: 10.2217/fon.09.128. Future Oncol. 2010. PMID: 20021212 Free PMC article. Review.
-
Neutrophils: game changers in glomerulonephritis?Trends Mol Med. 2010 Aug;16(8):368-78. doi: 10.1016/j.molmed.2010.06.002. Epub 2010 Jul 29. Trends Mol Med. 2010. PMID: 20667782 Free PMC article. Review.
References
-
- Murphy, P.M. 2002. International Union of Pharmacology. XXX. Update on chemokine receptor nomenclature. Pharmacol. Rev. 54:227–229. - PubMed
-
- Zlotnik, A., and O. Yoshie. 2000. Chemokines: a new classification system and their role in immunity. Immunity. 12:121–127. - PubMed
-
- Matsushima, K., K. Morishita, T. Yoshimura, S. Lavu, Y. Kobayashi, W. Lew, E. Appella, H.F. Kung, E.J. Leonard, and J.J. Oppenheim. 1988. Molecular cloning of a human monocyte-derived neutrophil chemotactic factor (MDNCF) and the induction of MDNCF mRNA by interleukin 1 and tumor necrosis factor. J. Exp. Med. 167:1883–1893. - PMC - PubMed
-
- Strieter, R.M., P.J. Polverini, S.L. Kunkel, D.A. Arenberg, M.D. Burdick, J. Kasper, J. Dzuiba, J. Van Damme, A. Walz, and D. Marriott. 1995. The functional role of the ELR motif in CXC chemokine-mediated angiogenesis. J. Biol. Chem. 270:27348–27357. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
