

Oncogenic bystander radiation effects in Patched heterozygous mouse cerebellum
- PMID: 18711141
- PMCID: PMC2517601
- DOI: 10.1073/pnas.0804186105
Oncogenic bystander radiation effects in Patched heterozygous mouse cerebellum
Abstract
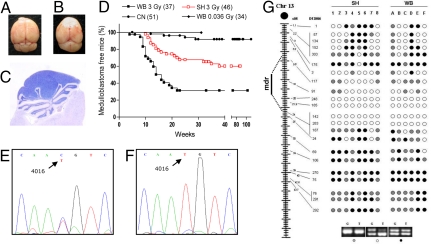
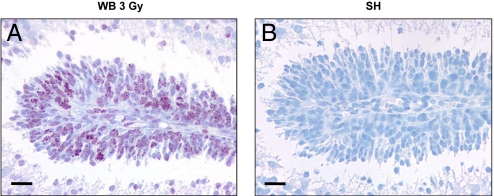
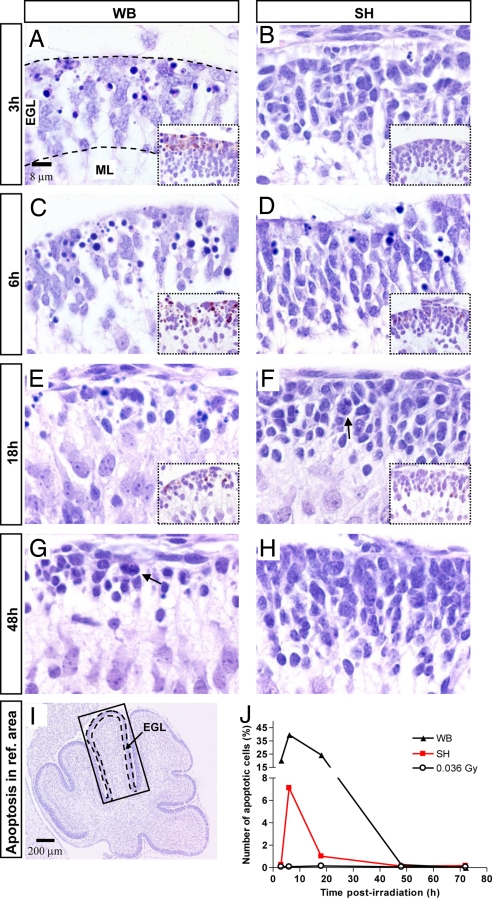
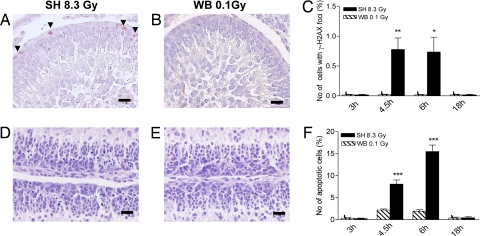
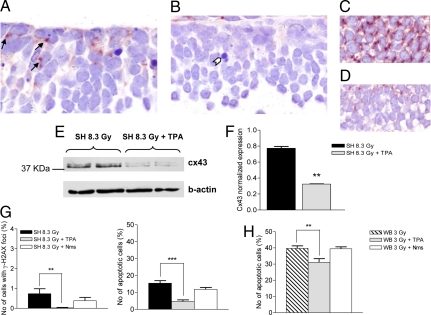
The central dogma of radiation biology, that biological effects of ionizing radiation are a direct consequence of DNA damage occurring in irradiated cells, has been challenged by observations that genetic/epigenetic changes occur in unexposed "bystander cells" neighboring directly-hit cells, due to cell-to-cell communication or soluble factors released by irradiated cells. To date, the vast majority of these effects are described in cell-culture systems, while in vivo validation and assessment of biological consequences within an organism remain uncertain. Here, we describe the neonatal mouse cerebellum as an accurate in vivo model to detect, quantify, and mechanistically dissect radiation-bystander responses. DNA double-strand breaks and apoptotic cell death were induced in bystander cerebellum in vivo. Accompanying these genetic events, we report bystander-related tumor induction in cerebellum of radiosensitive Patched-1 (Ptch1) heterozygous mice after x-ray exposure of the remainder of the body. We further show that genetic damage is a critical component of in vivo oncogenic bystander responses, and provide evidence supporting the role of gap-junctional intercellular communication (GJIC) in transmission of bystander signals in the central nervous system (CNS). These results represent the first proof-of-principle that bystander effects are factual in vivo events with carcinogenic potential, and implicate the need for re-evaluation of approaches currently used to estimate radiation-associated health risks.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- Little JB. Radiation carcinogenesis. Carcinogenesis. 2000;21:397–404. - PubMed
-
- Sowa M, Arthurs BJ, Estes BJ, Morgan WF. Effects of ionizing radiation on cellular structures, induced instability and carcinogenesis. EXS. 2006;96:293–301. - PubMed
-
- Nagasawa H, Little JB. Induction of sister chromatid exchanges by extremely low doses of alpha-particles. Cancer Res. 1992;52:6394–6396. - PubMed
-
- Azzam EI, de Toledo SM, Gooding T, Little JB. Intercellular communication is involved in the bystander regulation of gene expression in human cells exposed to very low fluences of alpha particles. Radiat Res. 1998;150:497–504. - PubMed
-
- Huang L, Kim PM, Nickoloff JA, Morgan WF. Targeted and nontargeted effects of low-dose ionizing radiation on delayed genomic instability in human cells. Cancer Res. 2007;67:1099–1104. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
