

Tilting at quixotic trait loci (QTL): an evolutionary perspective on genetic causation
- PMID: 18711218
- PMCID: PMC2516055
- DOI: 10.1534/genetics.108.094128
Tilting at quixotic trait loci (QTL): an evolutionary perspective on genetic causation
Abstract
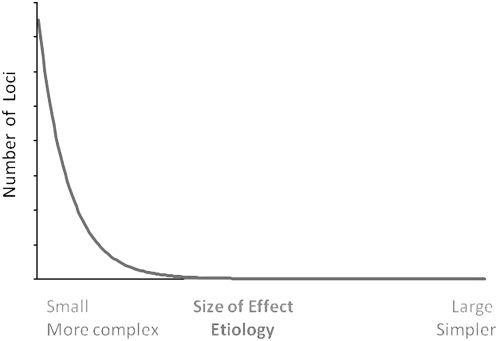
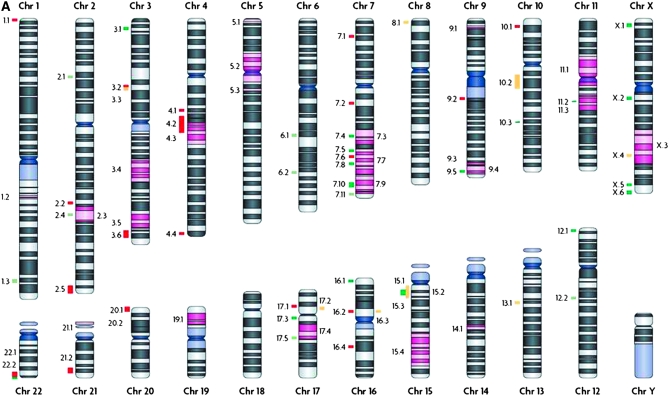
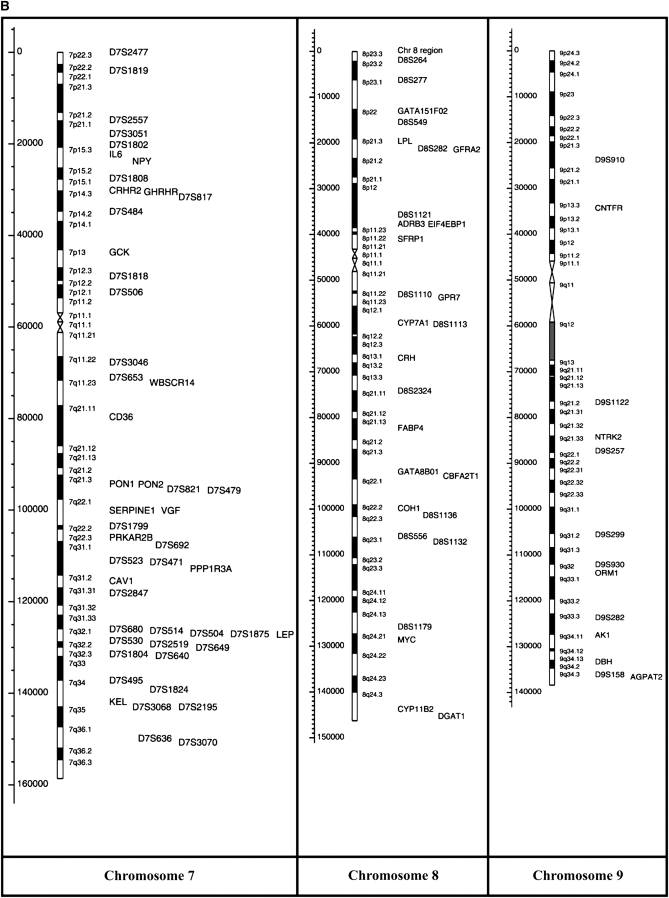
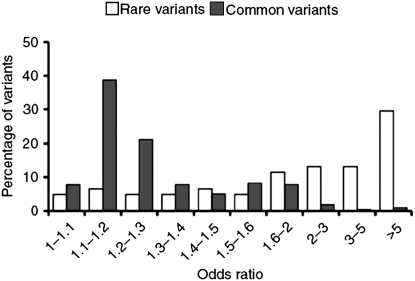
Recent years have seen great advances in generating and analyzing data to identify the genetic architecture of biological traits. Human disease has understandably received intense research focus, and the genes responsible for most Mendelian diseases have successfully been identified. However, the same advances have shown a consistent if less satisfying pattern, in which complex traits are affected by variation in large numbers of genes, most of which have individually minor or statistically elusive effects, leaving the bulk of genetic etiology unaccounted for. This pattern applies to diverse and unrelated traits, not just disease, in basically all species, and is consistent with evolutionary expectations, raising challenging questions about the best way to approach and understand biological complexity.
Figures

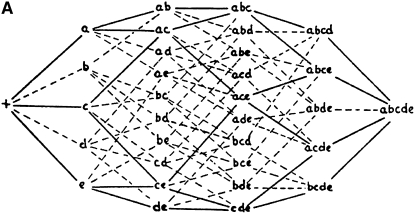
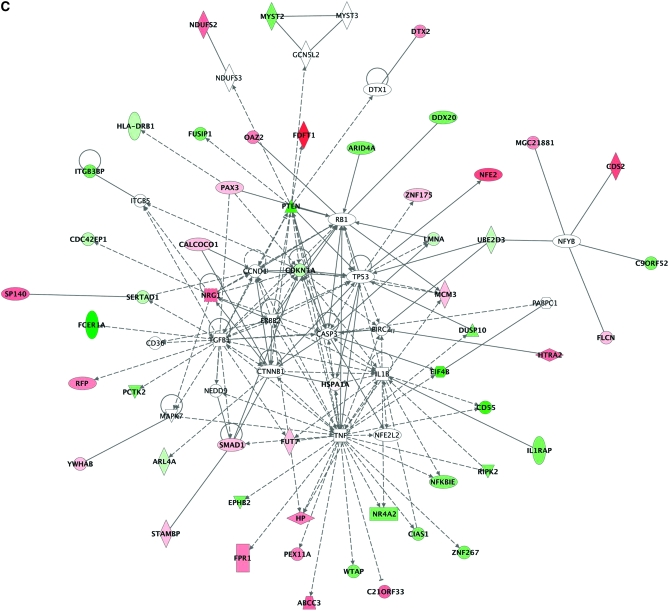
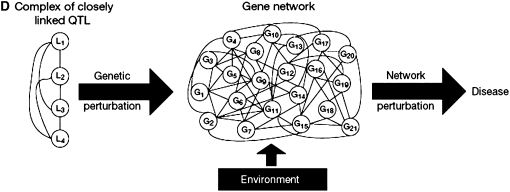
References
-
- Allen, N. C., S. Bagade, M. B. McQueen, J. P. Ioannidis, F. K. Kavvoura et al., 2008. Systematic meta-analyses and field synopsis of genetic association studies in schizophrenia: the SzGene database. Nat. Genet. 40 827–834. - PubMed
-
- Allikmets, R., and M. Dean, 2008. Bringing age-related macular degeneration into focus. Nat. Genet. 40 820–821. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
