

Evolution exacerbates the paradox of the plankton
- PMID: 18713864
- PMCID: PMC2527917
- DOI: 10.1073/pnas.0803032105
Evolution exacerbates the paradox of the plankton
Abstract
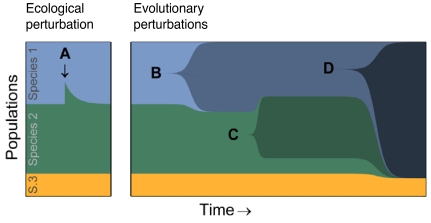
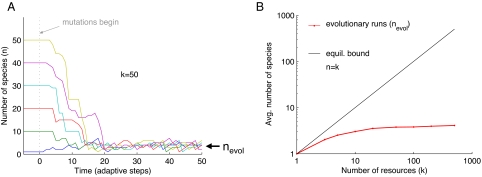
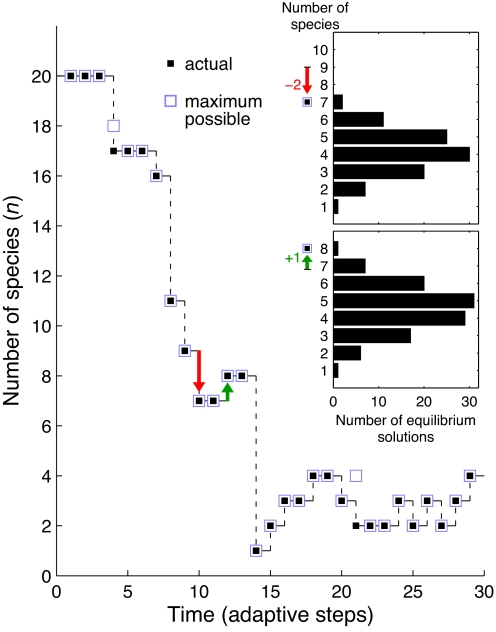
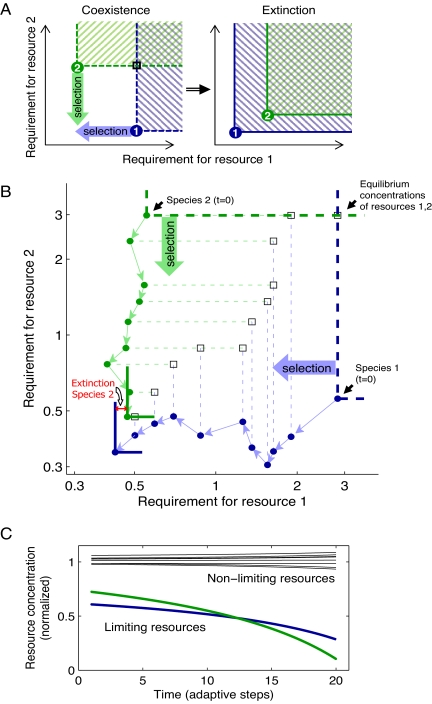
Can biodiversity evolve and persist in a uniform environment? This question is at the heart of the plankton paradox: in the natural world we observe many species sharing few resources, whereas the principle of competitive exclusion would lead us to expect that only a few species could coexist in such circumstances. To bridge the gap between theory and observation, previous studies have shown that the maximum number of species that can stably coexist is equal to the number of essential resources and that even more species can coexist out of equilibrium. These studies were viewed as a significant step toward a resolution of the paradox. Evolutionary dynamics, however, have been studied in this context only in limited cases, and it is largely unknown how mutations impact ecologically stable multispecies states, and whether large species consortia can spontaneously evolve. In the present study we introduce evolution to the standard ecological model of competition for essential resources. Combining numeric and analytic approaches, we find that ecologically stable species communities are severely destabilized by long-term evolutionary dynamics. Moreover, the number of species in spontaneously evolved consortia is much lower than the number of available resources. Contrary to expectations based on studies of two resources, these limits on biodiversity are not results of the occasional emergence of superspecies, superior to all competitors; nor are they alleviated by the inclusion of tradeoffs in resource utilization. Rather, we show that it is an accelerated depletion of limiting resources, combined with the essentiality of resources to all species, that leads invariably to catastrophic extinctions.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Hutchinson GE. The paradox of the plankton. Am Nat. 1961;95:137–145.
-
- Petersen R. The paradox of plankton: an equilibrium hypothesis. Am Nat. 1975;109:35–49.
-
- Monod J. La technique de culture continue theorie et applications. Ann Inst Pasteur. 1950;79:390–410.
-
- Herbert DR, Elsworth R, Telling RC. The continuous culture of bacteria: a theoretical and experimental study. J Gen Microbiol. 1956;14:601–622. - PubMed
-
- Dugdale RC. Nutrient limitation in the sea: Dynamics identification and significance. Limnol Oceanogr. 1967;12:685–695.
MeSH terms
LinkOut - more resources
Full Text Sources
Miscellaneous
