

Structural basis of Clostridium perfringens toxin complex formation
- PMID: 18716000
- PMCID: PMC2527888
- DOI: 10.1073/pnas.0803154105
Structural basis of Clostridium perfringens toxin complex formation
Abstract
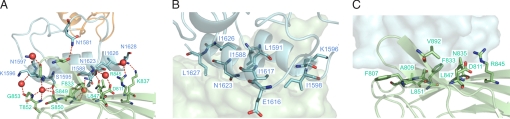
The virulent properties of the common human and livestock pathogen Clostridium perfringens are attributable to a formidable battery of toxins. Among these are a number of large and highly modular carbohydrate-active enzymes, including the mu-toxin and sialidases, whose catalytic properties are consistent with degradation of the mucosal layer of the human gut, glycosaminoglycans, and other cellular glycans found throughout the body. The conservation of noncatalytic ancillary modules among these enzymes suggests they make significant contributions to the overall functionality of the toxins. Here, we describe the structural basis of an ultra-tight interaction (K(a) = 1.44 x 10(11) M(-1)) between the X82 and dockerin modules, which are found throughout numerous C. perfringens carbohydrate-active enzymes. Extensive hydrogen-bonding and van der Waals contacts between the X82 and dockerin modules give rise to the observed high affinity. The mu-toxin dockerin module in this complex is positioned approximately 180 degrees relative to the orientation of the dockerin modules on the cohesin module surface within cellulolytic complexes. These observations represent a unique property of these clostridial toxins whereby they can associate into large, noncovalent multitoxin complexes that allow potentiation of the activities of the individual toxins by combining complementary toxin specificities.
Conflict of interest statement
The authors declare no conflict of interest.
Figures





References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
