

Increased blood pressure in mice lacking cytochrome P450 2J5
- PMID: 18716027
- PMCID: PMC2614614
- DOI: 10.1096/fj.08-114413
Increased blood pressure in mice lacking cytochrome P450 2J5
Abstract
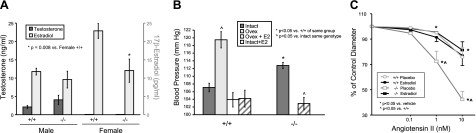
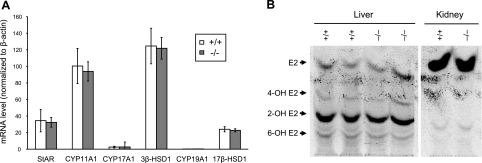
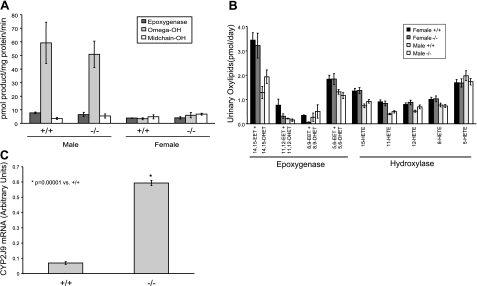
The cytochrome P450 (CYP) enzymes participate in a wide range of biochemical functions, including metabolism of arachidonic acid and steroid hormones. Mouse CYP2J5 is abundant in the kidney where its products, the cis-epoxyeicosatrienoic acids (EETs), modulate sodium transport and vascular tone. To define the physiological role of CYP2J5 in the kidney, knockout mice were generated using a conventional gene targeting approach. Cyp2j5 (-/-) mice develop normally and exhibit no overt renal pathology. While renal EET biosynthesis was apparently unaffected by the absence of CYP2J5, deficiency of this CYP in female mice was associated with increased blood pressure, enhanced proximal tubular transport rates, and exaggerated afferent arteriolar responses to angiotensin II and endothelin I. Interestingly, plasma 17beta-estradiol levels were reduced in female Cyp2j5 (-/-) mice and estrogen replacement restored blood pressure and vascular responsiveness to normal levels. There was no evidence of enhanced estrogen metabolism, or altered expression or activities of steroidogenic enzymes in female Cyp2j5 (-/-) mice, but their plasma levels of luteinizing hormone and follicle stimulating hormone were inappropriately low. Together, our findings illustrate a sex-specific role for CYP2J5 in regulation of blood pressure, proximal tubular transport, and afferent arteriolar responsiveness via an estrogen-dependent mechanism.
Figures
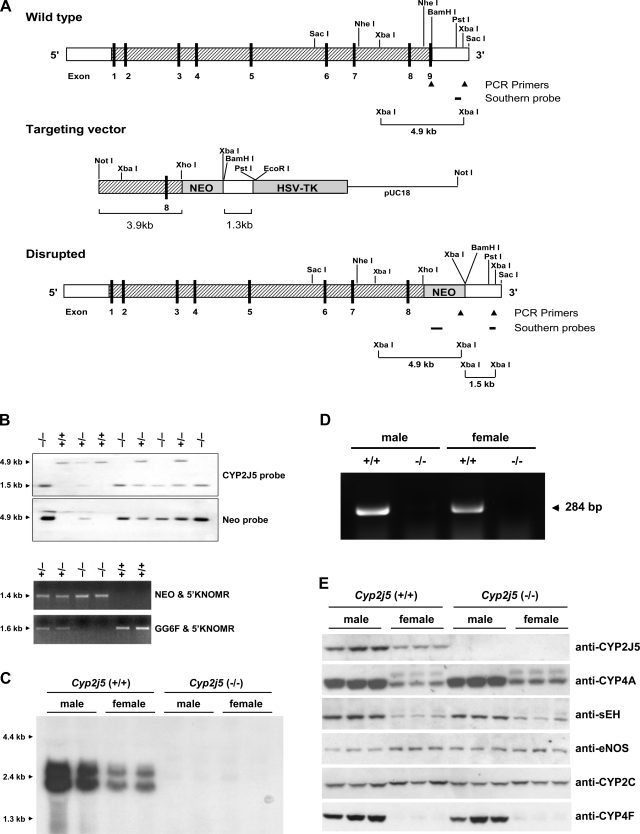
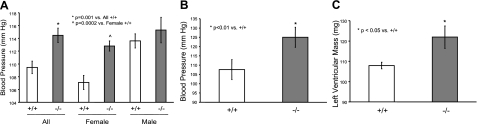
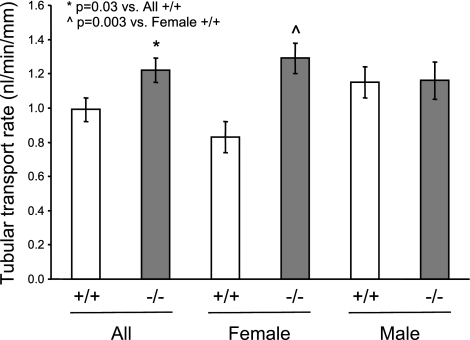
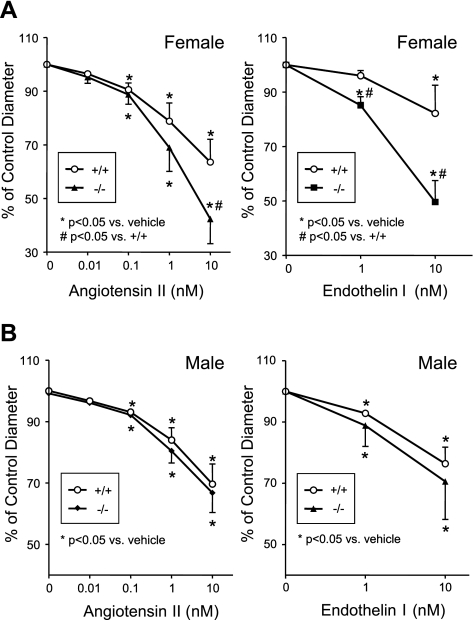
References
-
- Capdevila J H, Falck J R, Harris R C. Cytochrome P450 and arachidonic acid bioactivation: molecular and functional properties of the arachidonate monooxygenase. J Lipid Res. 2000;41:163–181. - PubMed
-
- Roman R J. P-450 metabolites of arachidonic acid in the control of cardiovascular function. Physiol Rev. 2002;82:131–185. - PubMed
-
- Zeldin D C. Epoxygenase pathways of arachidonic acid metabolism. J Biol Chem. 2001;276:36059–36062. - PubMed
-
- McGiff J C, Quilley J. 20-hydroxyeicosatetraenoic acid and epoxyeicosatrienoic acids and blood pressure. Curr Opin Nephrol Hypertens. 2001;10:231–237. - PubMed
-
- Imig J D, Zou A P, Stec D E, Harder D R, Falck J R, Roman R J. Formation and actions of 20-hydroxyeicosatetraenoic acid in rat renal arterioles. Am J Physiol. 1996;270:R217–227. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- MH39917/MH/NIMH NIH HHS/United States
- ES02710/ES/NIEHS NIH HHS/United States
- R01 HL059699/HL/NHLBI NIH HHS/United States
- R01 ES002710/ES/NIEHS NIH HHS/United States
- R01 MH039917/MH/NIMH NIH HHS/United States
- R01 DK069896/DK/NIDDK NIH HHS/United States
- Z01 ES025034/ImNIH/Intramural NIH HHS/United States
- DK38226/DK/NIDDK NIH HHS/United States
- R37 ES002710/ES/NIEHS NIH HHS/United States
- ES004699/ES/NIEHS NIH HHS/United States
- P01 DK038226/DK/NIDDK NIH HHS/United States
- HL6876/HL/NHLBI NIH HHS/United States
- P42 ES004699/ES/NIEHS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
