

Review
doi: 10.1016/j.bbagrm.2008.07.008.
Epub 2008 Jul 30.
Protein modifications in transcription elongation
Affiliations
- PMID: 18718879
- PMCID: PMC2641038
- DOI: 10.1016/j.bbagrm.2008.07.008
Item in Clipboard
Review
Protein modifications in transcription elongation
Biochim Biophys Acta.
2009 Jan.
Abstract
Posttranslational modifications (PTMs) of proteins play essential roles in regulating signaling, protein-protein modifications and subcellular localization. In this review, we focus on posttranslational modification of histones and RNA polymerase II (RNAPII) and their roles in gene transcription. A survey of the basic features of PTMs is provided followed by a more detailed account of how PTMs on histones and RNAPII regulate transcription in the model organism Saccharomyces cerevisiae. We emphasize the interconnections between histone and RNAPII PTMs and speculate upon the larger role PTMs have in regulating protein function in the cell.
Figures
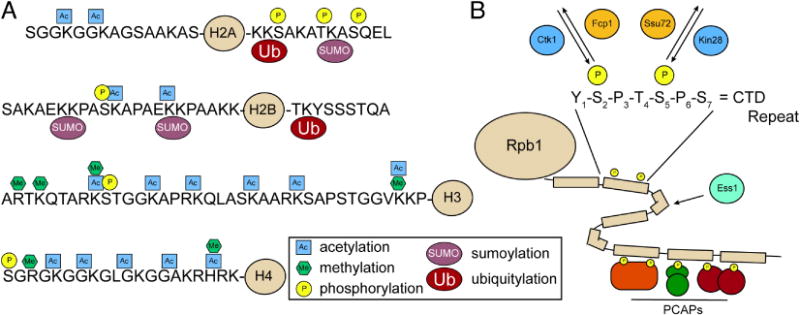
Posttranslational modifications (PTMs) found on histones and RNA polymerase II (RNAPII). (A) Most common PTMs found on the N- and C-terminal tails of the four canonical histones in yeast. (B) Sites of phosphorylation within the 26-repeat C-terminal domain (CTD) of Rpb1, the largest subunit of RNAPII. Phosphorylation of Ser5 is primarily catalyzed by Kin28 and removed by Ssu72. Phosphorylation of Ser2 is primarily catalyzed by Ctk1 and removed by Fcp1. Ess1 is thought to facilitate prolyl isomerization within the CTD. Differential phosphorylation of the CTD leads to recruitment of distinct phosphor-CTD associating proteins (PCAPs) in a spatially and temporally controlled manner to regulate specific transcription-associated processes (e.g. chromatin remodeling and mRNA processing).
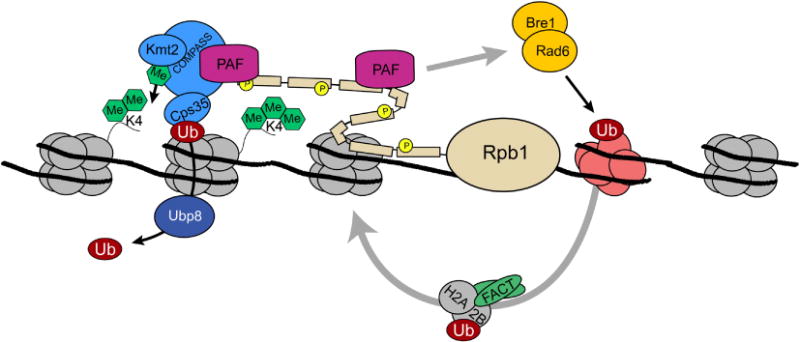
H3K4 methylation and transcriptional activation. The COMPASS methyltransferase is recruited to the promoter regions of genes through interactions with the PAF complex and serine 5 phosphorylation, as well as by making contacts with H2Bub1 through Cps35 (See text). Recruitment of COMPASS leads to the restricted establishment of H3K4me3 near the transcriptional start site This modification may serve to stabilize PIC formation and recruit chromatin remodeling and histone modifying activities that facilitate the transcription process (see text for details). Also associated with COMPASS and the elongating RNAPII complex is the histone chaperone complex, FACT, which disassembles and reassembles nucleosomes to promote transcription. FACT function and COMPASS activity are both functionally regulated, at least in part, through H2Bub1 [110].
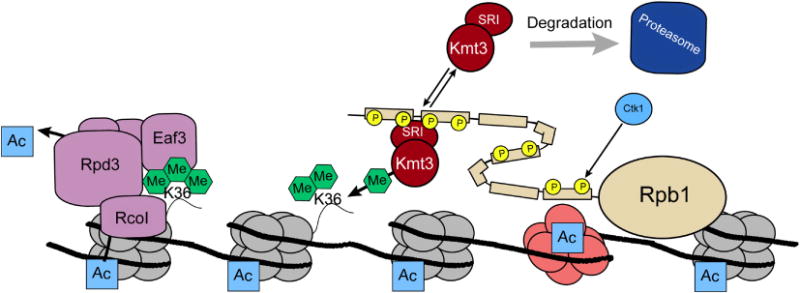
Role of H3K36 methylation in transcriptional dampening. Following CTD phosphorylation by Ctk1, the Kmt3(Set2) methyltransferase is recruited to the elongating form of RNAPII via its SRI domain [171]. Kmt3(Set2) methylates H3K36, leading to Rpd3(S) deacetylase recruitment and deacetylation of ORF specific nucleosomes. This event regulates the removal of elongation-linked acetylation and thereby prevents inappropriate initiation events from occurring in gene bodies. Thus, H3K36me serves to maintain proper transcription initiation at the promoter regions of genes.
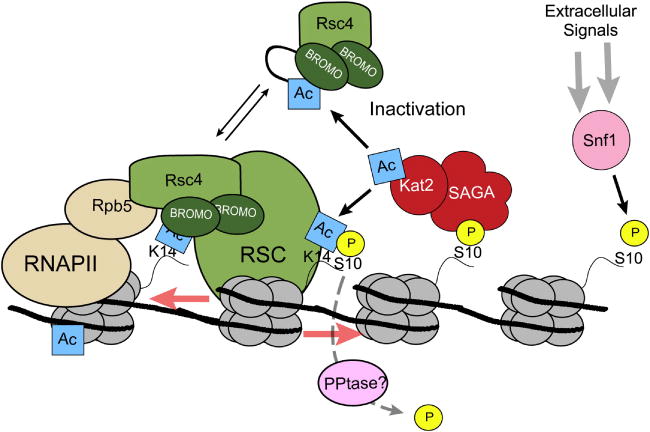
Acetylation both stimulates and inhibits RSC chromatin remodeling activities. Kat2(Gcn5) is recruited to acetylate H3K14 by Snf1-mediated phosphorylation of H3S10. During transcription, RSC chromatin remodeling activities is recruited via Rsc4 which binds to H3K14ac via its tandem bromodomains and with RNAPII through interactions with Rpb5. Rsc4 can also be directly acetylated by Kat2(Gcn5) leading to inactivation of the complex.
References
-
- Black DL. Mechanisms of alternative pre-messenger RNA splicing. Annu Rev Biochem. 2003;72:291–336. - PubMed
-
- Maniatis T, Tasic B. Alternative pre-mRNA splicing and proteome expansion in metazoans. Nature. 2002;418:236–243. - PubMed
-
- Uy R, Wold F. Posttranslational covalent modification of proteins. Science. 1977;198:890–896. - PubMed
-
- Walsh CT. Posttranslational modifications of proteins: expanding nature's inventory. Roberts and Co.; Englewood, CO: 2006.
-
- Dunlop PC, Bodley JW. Biosynthetic labeling of diphthamide in Saccharomyces cerevisiae. J Biol Chem. 1983;258:4754–4758. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
