

Inactivation of median preoptic nucleus causes c-Fos expression in hypocretin- and serotonin-containing neurons in anesthetized rat
- PMID: 18722360
- PMCID: PMC2592253
- DOI: 10.1016/j.brainres.2008.07.115
Inactivation of median preoptic nucleus causes c-Fos expression in hypocretin- and serotonin-containing neurons in anesthetized rat
Abstract
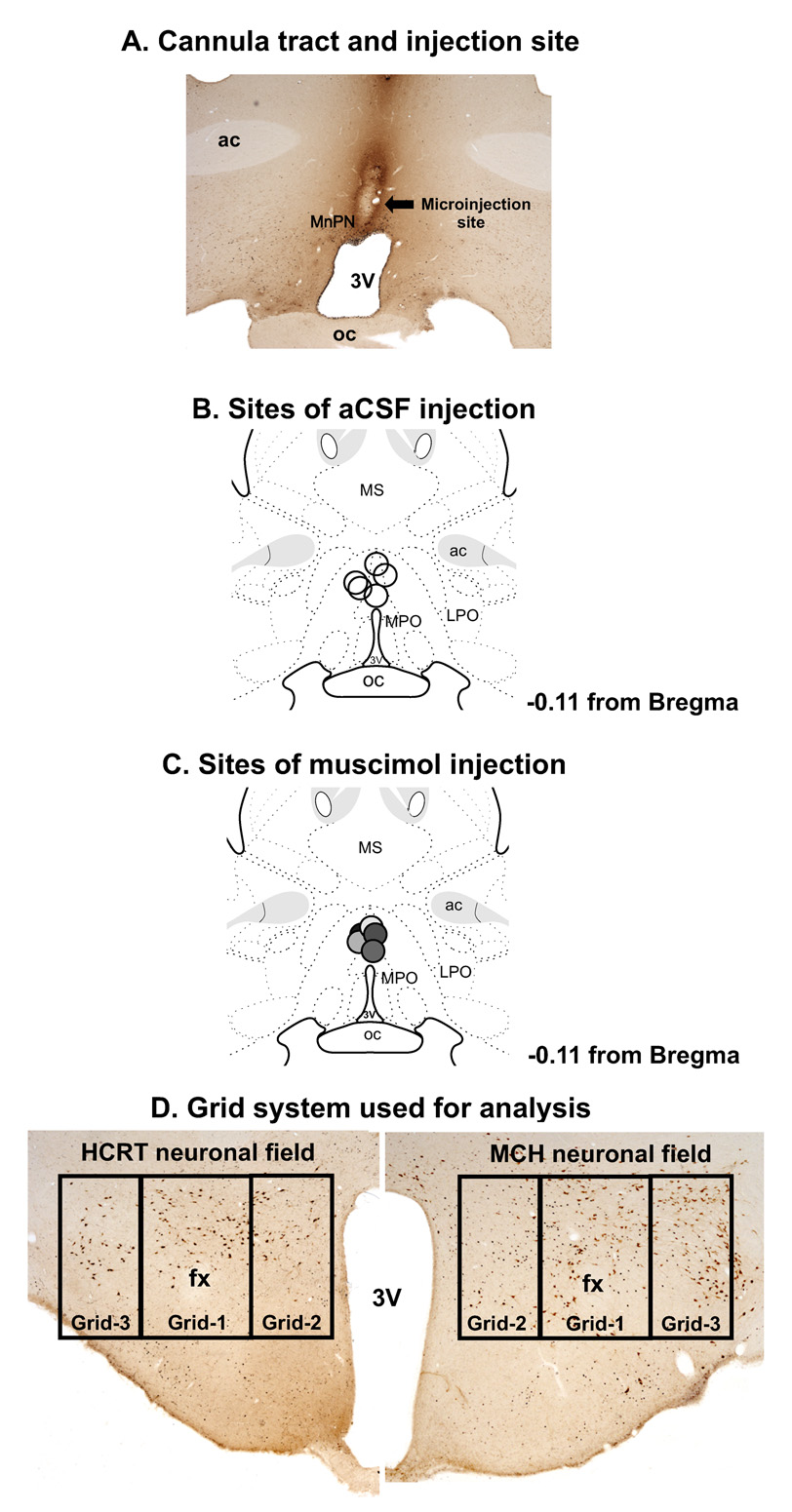
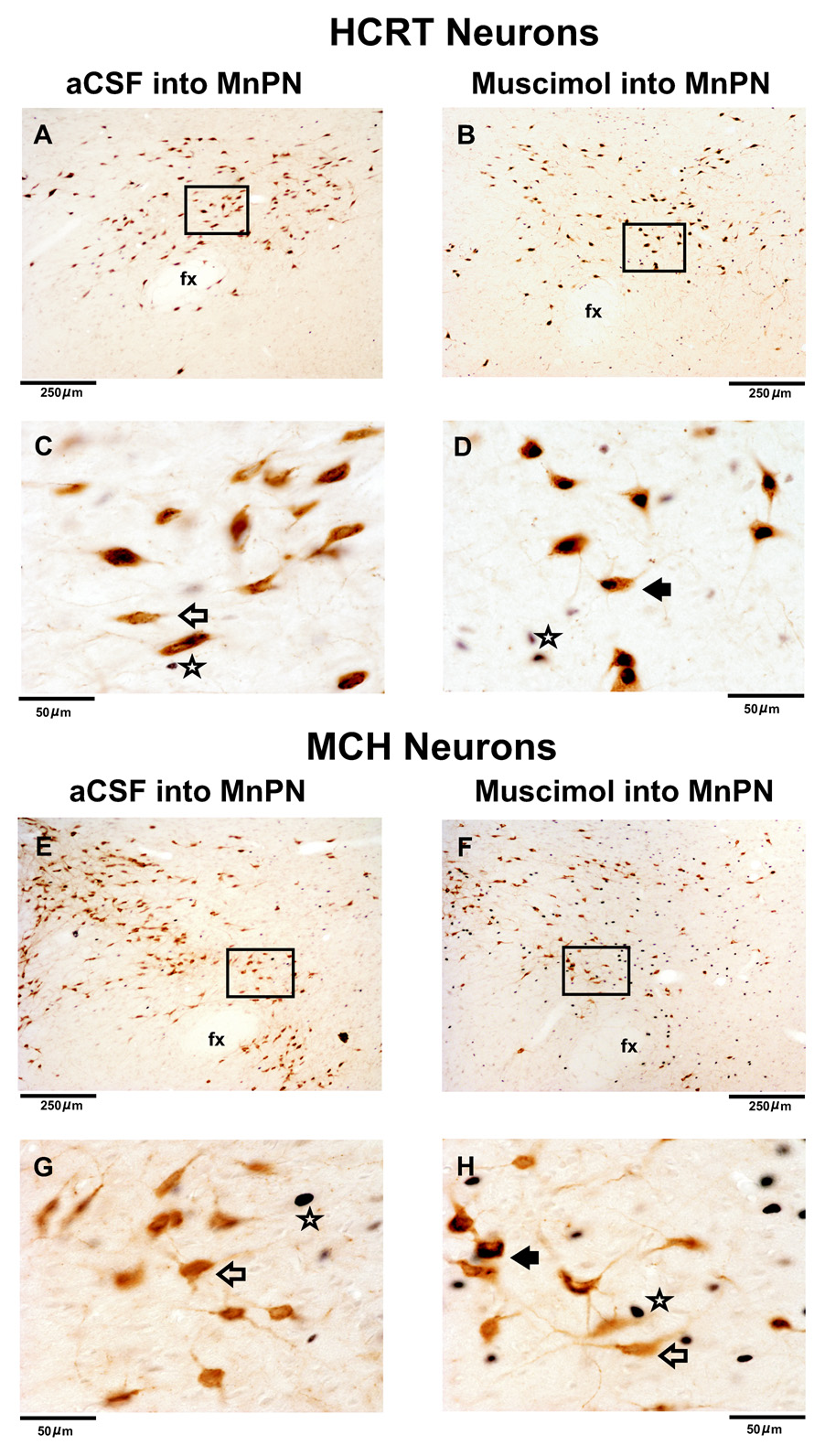
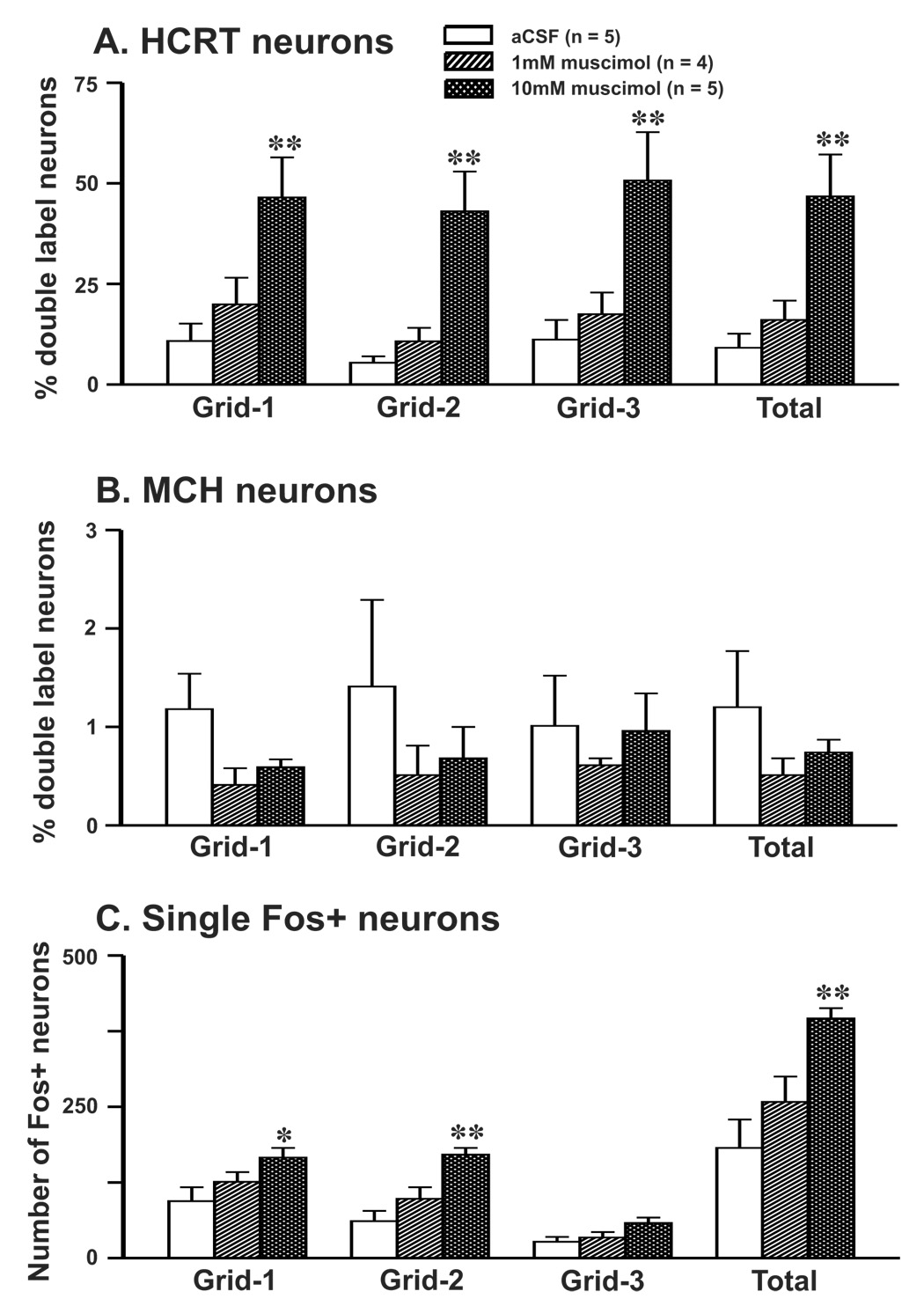
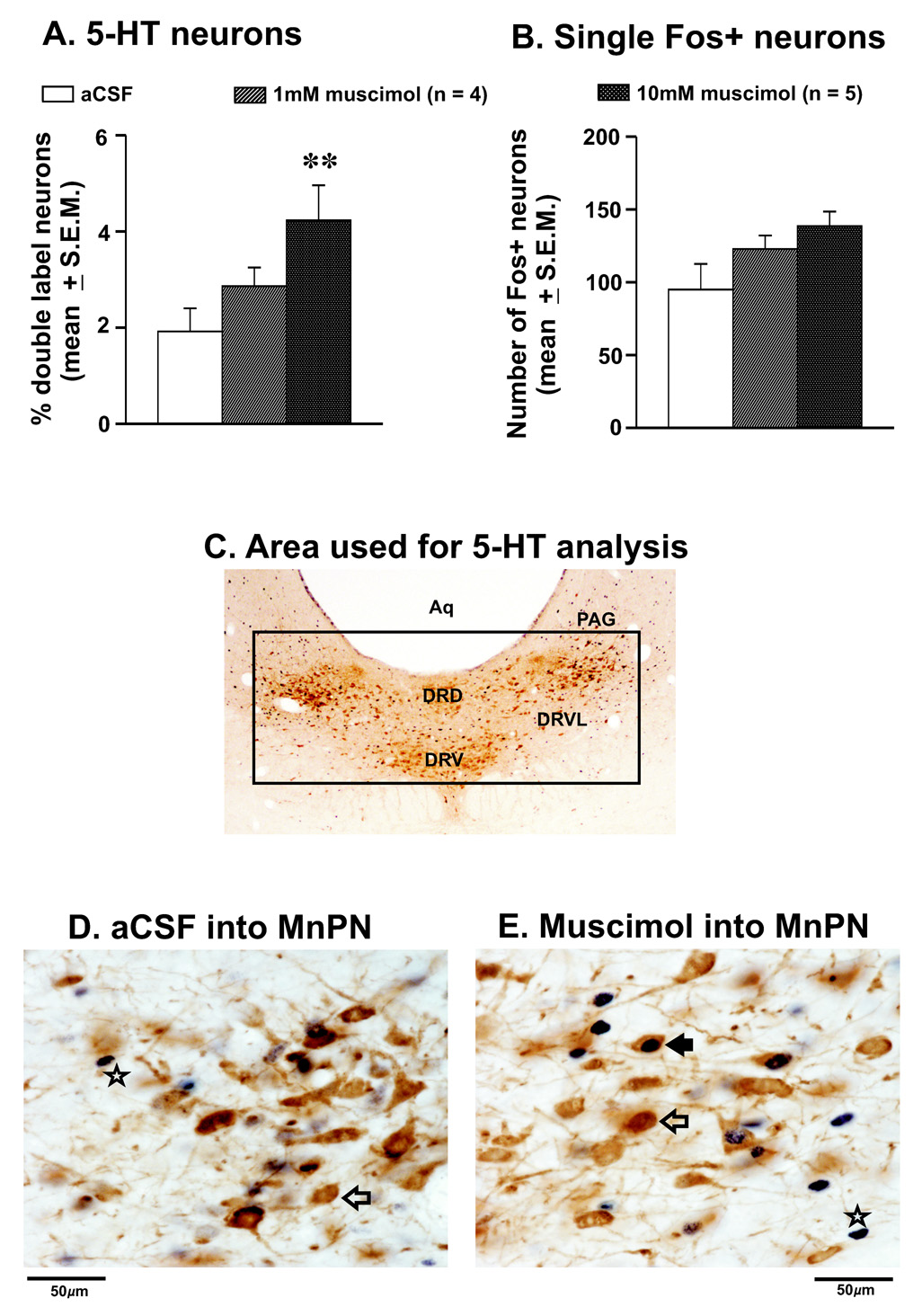
The median preoptic nucleus (MnPN) of the hypothalamus contains sleep-active neurons including sleep-active GABAergic neurons and is involved in the regulation of nonREM/REM sleep. The hypocretinergic (HCRT) neurons of the perifornical-lateral hypothalamic area (PF-LHA) and serotonergic (5-HT) neurons of the dorsal raphe nucleus (DRN) are mostly active during waking and have been implicated in the regulation of arousal. MnPN GABAergic neurons project to the PF-LHA and DRN. It is hypothesized that MnPN promotes sleep by inhibiting multiple arousal systems including HCRT and other wake-active neurons within the PF-LHA and 5-HT neurons in the DRN. We examined the effects of inactivation of MnPN neurons by locally microinjecting 0.2 microl of 1 mM or 10 mM solutions of a GABA(A) receptor agonist, muscimol, into the MnPN on Fos expression (Fos-IR) in the PF-LHA neurons including HCRT neurons and 5-HT neurons in the DRN in anesthetized rats. Compared to artificial cerebrospinal fluid control, microinjection of muscimol into the MnPN resulted in significantly higher percentages of HCRT and non-HCRT neurons in the PF-LHA and 5-HT neurons in the DRN that exhibited Fos-IR. The percentage of melanin-concentrating hormone (MCH)+/Fos+ neurons in the PF-LHA did not change after muscimol treatments. These results support a hypothesis that the activation of MnPN neurons contributes to the suppression of wake-promoting systems including HCRT and other unidentified neurons in the PF-LHA and 5-HT neurons in the DRN. These results also suggest that MCH neurons may not be under MnPN inhibitory control. These findings are consistent with a hypothesized role of MnPN in sleep regulation.
Figures
Similar articles
-
GABA-mediated control of hypocretin- but not melanin-concentrating hormone-immunoreactive neurones during sleep in rats.J Physiol. 2005 Mar 1;563(Pt 2):569-82. doi: 10.1113/jphysiol.2004.076927. Epub 2004 Dec 21. J Physiol. 2005. PMID: 15613374 Free PMC article.
-
Effects of serotonin on perifornical-lateral hypothalamic area neurons in rat.Eur J Neurosci. 2007 Jan;25(1):201-12. doi: 10.1111/j.1460-9568.2006.05268.x. Eur J Neurosci. 2007. PMID: 17241281
-
The median preoptic nucleus reciprocally modulates activity of arousal-related and sleep-related neurons in the perifornical lateral hypothalamus.J Neurosci. 2007 Feb 14;27(7):1616-30. doi: 10.1523/JNEUROSCI.3498-06.2007. J Neurosci. 2007. PMID: 17301170 Free PMC article.
-
The role of dorsal raphe nucleus serotonergic and non-serotonergic neurons, and of their receptors, in regulating waking and rapid eye movement (REM) sleep.Sleep Med Rev. 2010 Oct;14(5):319-27. doi: 10.1016/j.smrv.2009.10.003. Epub 2010 Feb 12. Sleep Med Rev. 2010. PMID: 20153670 Review.
-
Physiological arousal: a role for hypothalamic systems.Cell Mol Life Sci. 2008 May;65(10):1475-88. doi: 10.1007/s00018-008-7521-8. Cell Mol Life Sci. 2008. PMID: 18351292 Free PMC article. Review.
Cited by
-
Distinct patterns of neuronal inputs and outputs of the juxtaparaventricular and suprafornical regions of the lateral hypothalamic area in the male rat.Brain Res Rev. 2010 Sep;64(1):14-103. doi: 10.1016/j.brainresrev.2010.02.002. Epub 2010 Feb 17. Brain Res Rev. 2010. PMID: 20170674 Free PMC article. Review.
-
Adenosine A(2A) receptors regulate the activity of sleep regulatory GABAergic neurons in the preoptic hypothalamus.Am J Physiol Regul Integr Comp Physiol. 2013 Jul 1;305(1):R31-41. doi: 10.1152/ajpregu.00402.2012. Epub 2013 May 1. Am J Physiol Regul Integr Comp Physiol. 2013. PMID: 23637137 Free PMC article.
-
Characteristics of sleep-active neurons in the medullary parafacial zone in rats.Sleep. 2018 Oct 1;41(10):zsy130. doi: 10.1093/sleep/zsy130. Sleep. 2018. PMID: 29986116 Free PMC article.
-
c-Fos expression in neurons projecting from the preoptic and lateral hypothalamic areas to the ventrolateral periaqueductal gray in relation to sleep states.Neuroscience. 2011 Aug 11;188:55-67. doi: 10.1016/j.neuroscience.2011.05.016. Epub 2011 May 17. Neuroscience. 2011. PMID: 21601616 Free PMC article.
-
Preoptic Area Modulation of Arousal in Natural and Drug Induced Unconscious States.Front Neurosci. 2021 Feb 12;15:644330. doi: 10.3389/fnins.2021.644330. eCollection 2021. Front Neurosci. 2021. PMID: 33642991 Free PMC article. Review.
References
-
- Abrahamson EE, Moore RY. The posterior hypothalamic area: chemoarchitecture and afferent connections. Brain Res. 2001;889:1–22. - PubMed
-
- Auerbach SB, Minzenberg MJ, Wilkinson LO. Extracellular serotonin and 5-hydroxyindoleacetic acid in hypothalamus of the unanesthetized rat measured by in vivo dialysis coupled to high-performance liquid chromatography with electrochemical detection: dialysate serotonin reflects neuronal release. Brain Res. 1989;499:281–290. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
