

Histone modifying enzymes: structures, mechanisms, and specificities
- PMID: 18722564
- PMCID: PMC4059211
- DOI: 10.1016/j.bbagrm.2008.07.009
Histone modifying enzymes: structures, mechanisms, and specificities
Abstract
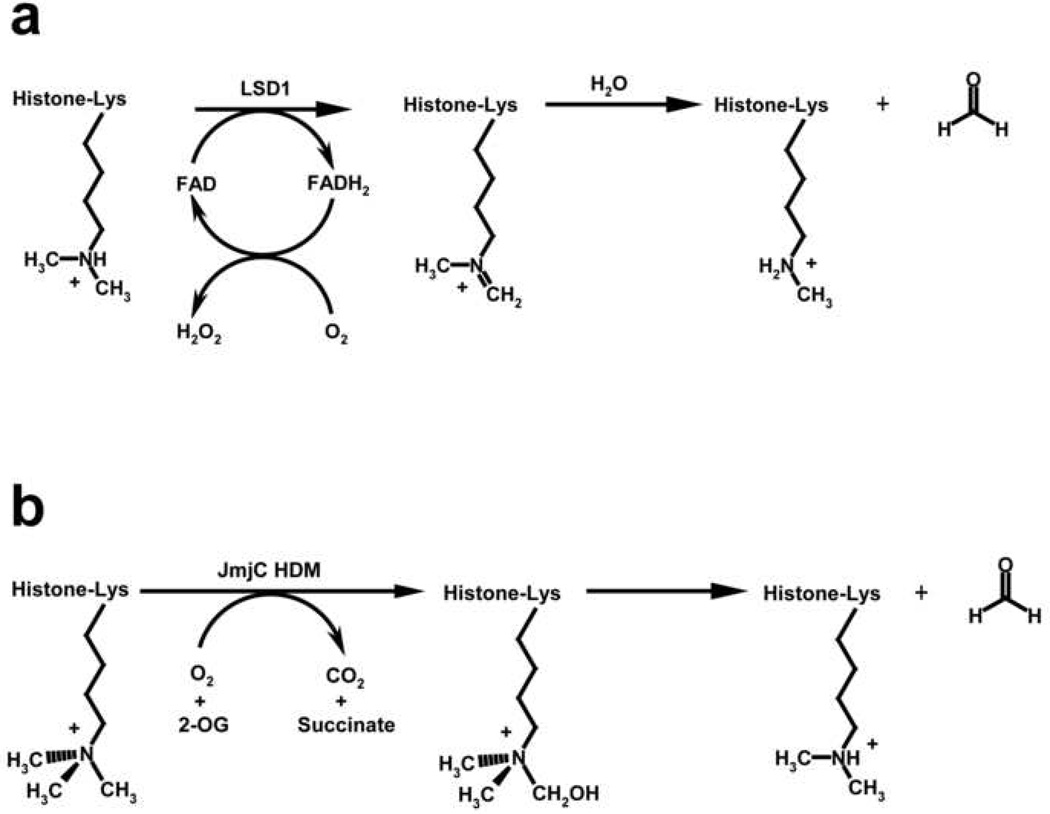
Histone modifying enzymes catalyze the addition or removal of an array of covalent modifications in histone and non-histone proteins. Within the context of chromatin, these modifications regulate gene expression as well as other genomic functions and have been implicated in establishing and maintaining a heritable epigenetic code that contributes to defining cell identity and fate. Biochemical and structural characterization of histone modifying enzymes has yielded important insights into their respective catalytic mechanisms, substrate specificities, and regulation. In this review, we summarize recent advances in understanding these enzymes, highlighting studies of the histone acetyltransferases (HATs) p300 (also now known as KAT3B) and Rtt109 (KAT11) and the histone lysine demethylases (HDMs) LSD1 (KDM1) and JMJD2A (KDM4A), present overriding themes that derive from these studies, and pose remaining questions concerning their regulatory roles in mediating DNA transactions.
Figures






References
-
- Luger K, Hansen JC. Nucleosome and chromatin fiber dynamics. Curr Opin Struct Biol. 2005;15:188–196. - PubMed
-
- Kouzarides T. Chromatin modifications and their function. Cell. 2007;128:693–705. - PubMed
-
- Murray K. The Occurrence of Epsilon-N-Methyl Lysine in Histones. Biochemistry. 1964;3:10–15. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
