

Review
doi: 10.1016/j.mrfmmm.2008.07.011.
Epub 2008 Aug 3.
Epigenetic mechanisms of facioscapulohumeral muscular dystrophy
Affiliations
- PMID: 18723032
- PMCID: PMC2650037
- DOI: 10.1016/j.mrfmmm.2008.07.011
Item in Clipboard
Review
Epigenetic mechanisms of facioscapulohumeral muscular dystrophy
Mutat Res.
.
Abstract
Facioscapulohumeral muscular dystrophy (FSHD) seems to be caused by a complex epigenetic disease mechanism as a result of contraction of the polymorphic macrosatellite repeat D4Z4 on chromosome 4qter. Currently, the exact mechanism causing the FSHD phenotype is still not elucidated. In this review, we discuss the genetic and epigenetic changes observed in patients with FSHD and the possible disease mechanisms that may be associated with FSHD pathogenesis.
Conflict of interest statement
The authors declare that there are no conflicts of interest.
Figures
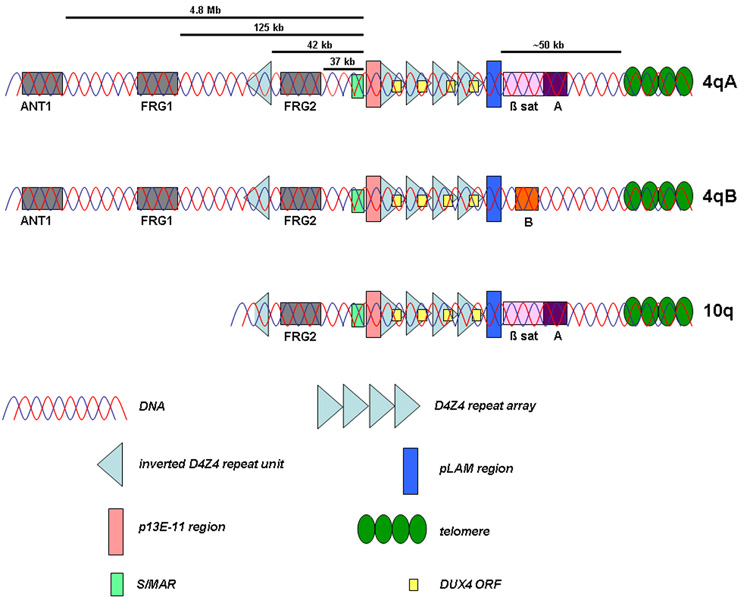
The subtelomere of chromosome 10q contains a repeat array that is highly homologous to D4Z4 on 4qter. The homology extends both in proximal (~40 kb upstream) and distal direction. In addition, two allelic variants of the 4q subtelomere have been identified. The presence of beta satellite DNA distal to D4Z4 on 4qA-type alleles is the most prominent difference between these allelic variants.
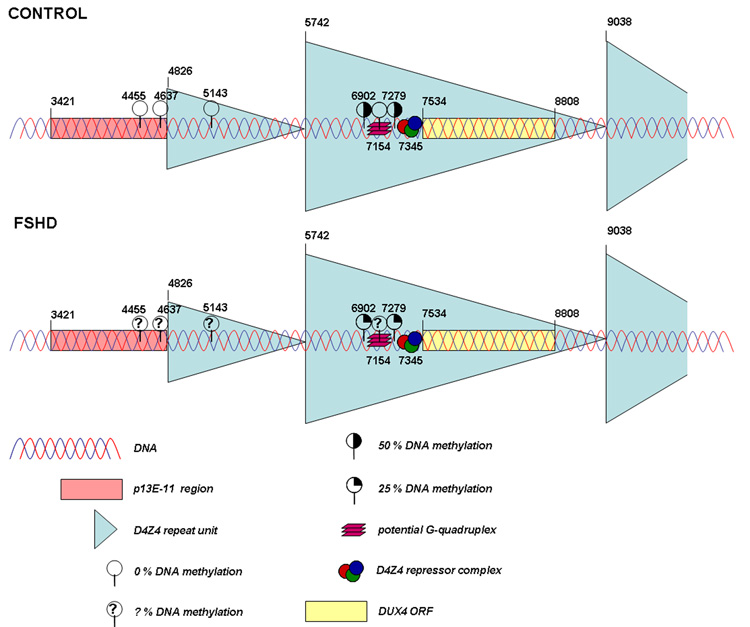
In control individuals methylation levels of 50 % are identified at two CpG dinucleotides while in patients with FSHD significant hypomethylation is found at these sites (25 % methylation). Interestingly, part of the proximal repeat unit seems to be resistant to DNA methylation (0 % methylation) and is more accessible to DNaseI in cancer tissues, suggesting the presence of a boundary element at the junction of D4Z4 and p13E-11. Finally, a subregion within each D4Z4 repeat unit that may form a G-quadruplex also shows resistance to cancer-linked hypermethylation (0 % methylation). In somatic control DNA samples, similar results were obtained, although at a lower intensity. FSHD DNA samples have not yet been tested (? % DNA methylation). The presented nucleotide positions are based on AF117653 (GenBank).
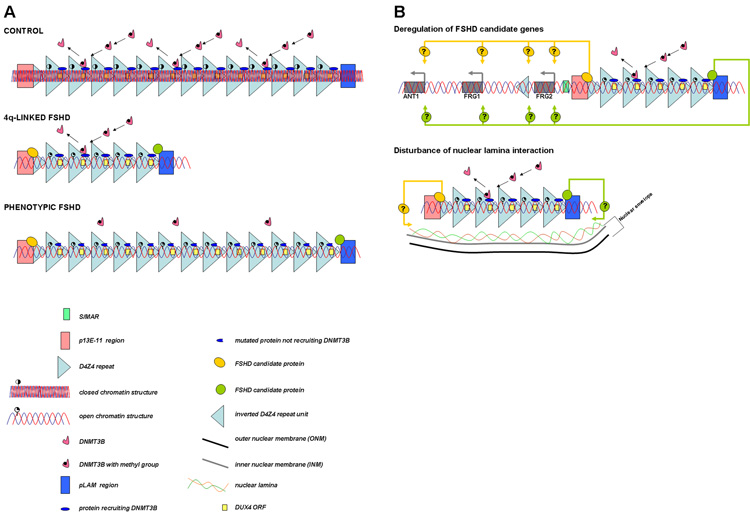
A. In controls the D4Z4 repeat array is packed as facultative heterochromatin. In patients with 4q-linked FSHD only below 11 D4Z4 repeat units, a more open chromatin structure is induced. Below this threshold an insufficient level of a protein (complex) responsible for D4Z4 methylation or a presently unidentified chromatin modifier (complex) may be present at D4Z4. In patients with phenotypic FSHD, changes in chromatin structure may occur because of a yet unidentified gene defect responsible for the epigenetic modifications at D4Z4. Therefore, an open chromatin structure is present in these patients even above the critical threshold of ten D4Z4 repeat units. Because of the open chromatin structure in patients with FSHD, binding of protein(s) to D4Z4 that normally do not bind may occur. This will most likely occur either proximal or distal to D4Z4 or to a critical D4Z4 element, since the amount of repeat units differs significantly between 4q-linked and phenotypic FSHD, while their phenotypes are highly similar. In addition, specific SNPs discriminating between the different haplotypes may be important for the binding of these proteins. B. When the chromatin structure is in a more open conformation, candidate genes may be deregulated in cis (upper panel) and the interaction with the nuclear envelope may be disturbed (lower panel).
References
-
- Padberg GW. Facioscapulohumeral disease, PhD thesis. Leiden: Leiden University; 1982.
-
- Wijmenga C, Frants RR, Brouwer OF, Moerer P, Weber JL, Padberg GW. Location of facioscapulohumeral muscular dystrophy gene on chromosome 4. Lancet. 1990;336:651–653. - PubMed
-
- van Deutekom JC, Wijmenga C, van Tienhoven EA, Gruter AM, Hewitt JE, Padberg GW, van Ommen GJ, Hofker MH, Frants RR. FSHD associated DNA rearrangements are due to deletions of integral copies of a 3.2 kb tandemly repeated unit. Hum. Mol. Genet. 1993;2:2037–2042. - PubMed
-
- Wijmenga C, Hewitt JE, Sandkuijl LA, Clark LN, Wright TJ, Dauwerse HG, Gruter AM, Hofker MH, Moerer P, Williamson R. Chromosome 4q DNA rearrangements associated with facioscapulohumeral muscular dystrophy. Nat.Genet. 1992;2:26–30. - PubMed
-
- Lunt PW, Jardine PE, Koch MC, Maynard J, Osborn M, Williams M, Harper PS, Upadhyaya M. Correlation between fragment size at D4F104S1 and age at onset or at wheelchair use, with a possible generational effect, accounts for much phenotypic variation in 4q35-facioscapulohumeral muscular dystrophy (FSHD) Hum.Mol.Genet. 1995;4:951–958. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
