

Hepatitis C viral NS3-4A protease activity is enhanced by the NS3 helicase
- PMID: 18723512
- PMCID: PMC2573085
- DOI: 10.1074/jbc.M804065200
Hepatitis C viral NS3-4A protease activity is enhanced by the NS3 helicase
Abstract
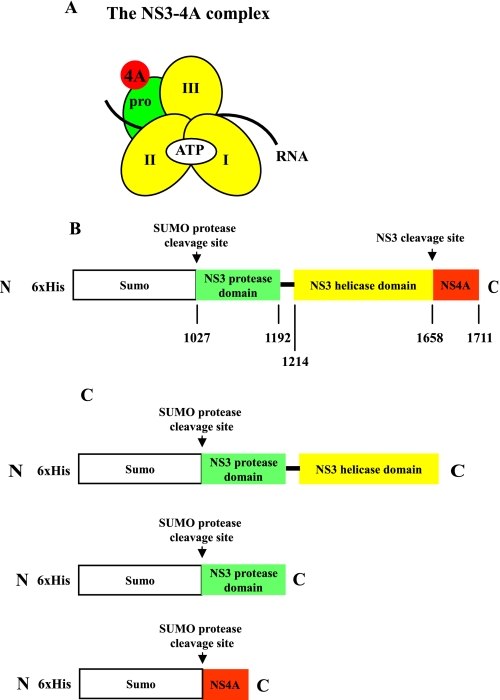
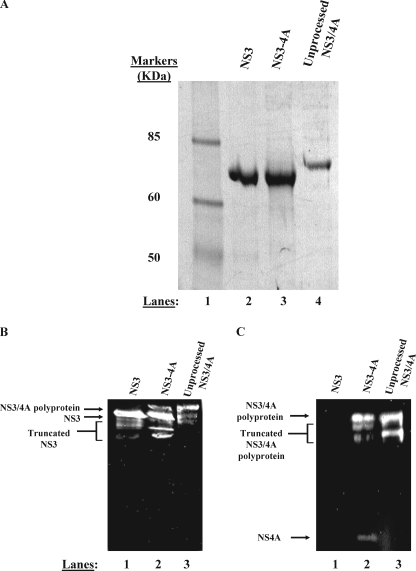
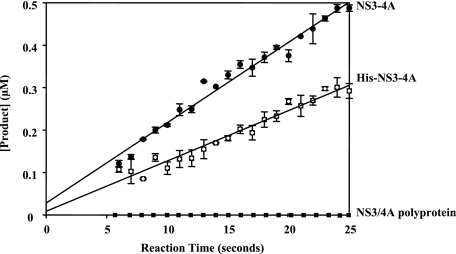
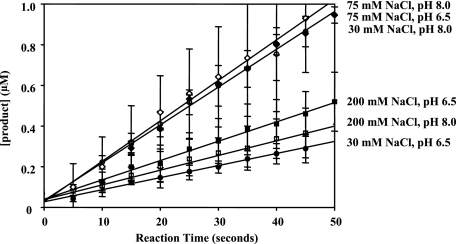
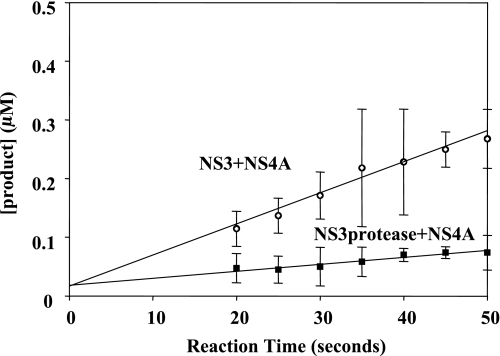
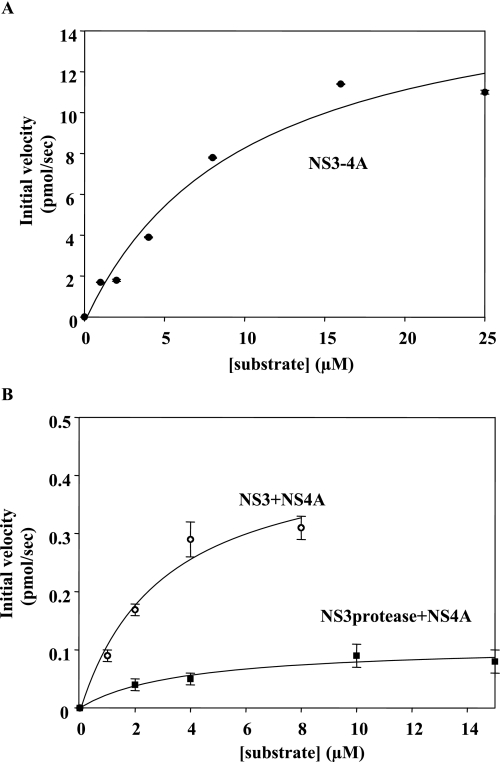
Non-structural protein 3 (NS3) is a multifunctional enzyme possessing serine protease, NTPase, and RNA unwinding activities that are required for hepatitis C viral (HCV) replication. HCV non-structural protein 4A (NS4A) binds to the N-terminal NS3 protease domain to stimulate NS3 serine protease activity. In addition, the NS3 protease domain enhances the RNA binding, ATPase, and RNA unwinding activities of the C-terminal NS3 helicase domain (NS3hel). To determine whether NS3hel enhances the NS3 serine protease activity, we purified truncated and full-length NS3-4A complexes and examined their serine protease activities under a variety of salt and pH conditions. Our results indicate that the helicase domain enhances serine protease activity, just as the protease domain enhances helicase activity. Thus, the two enzymatic domains of NS3-4A are highly interdependent. This is the first time that such a complete interdependence has been demonstrated for a multifunctional, single chain enzyme. NS3-4A domain interdependence has important implications for function during the viral lifecycle as well as for the design of inhibitor screens that target the NS3-4A protease.
Figures
References
-
- Lohmann, V., Korner, F., Koch, J., Herian, U., Theilmann, L., and Bartenschlager, R. (1999) Science 285 110-113 - PubMed
-
- de la Cruz, J., Kressler, D., and Linder, P. (1999) Trends Biochem. Sci. 24 192-198 - PubMed
-
- Taliani, M., Bianchi, E., Narjes, F., Fossatelli, M., Urbani, A., Steinkuhler, C., De Francesco, R., and Pessi, A. (1996) Anal. Biochem. 240 60-67 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
