

Beta-catenin levels influence rapid mechanical responses in osteoblasts
- PMID: 18723514
- PMCID: PMC2570859
- DOI: 10.1074/jbc.M801907200
Beta-catenin levels influence rapid mechanical responses in osteoblasts
Abstract
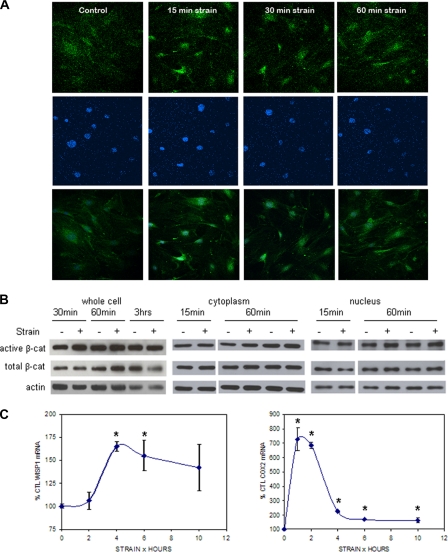
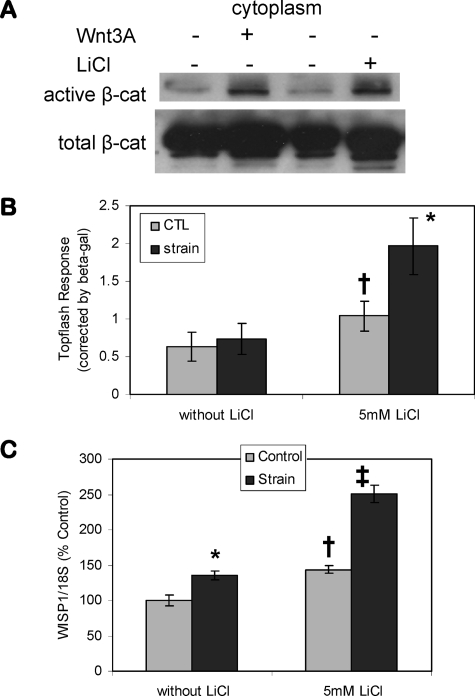
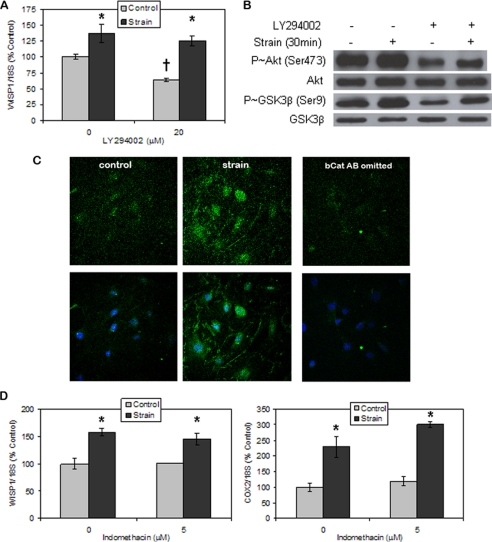
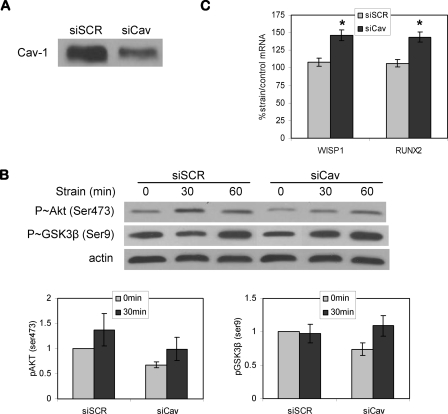
Mechanical loading of bone initiates an anabolic, anticatabolic pattern of response, yet the molecular events involved in mechanical signal transduction are not well understood. Wnt/beta-catenin signaling has been recognized in promoting bone anabolism, and application of strain has been shown to induce beta-catenin activation. In this work, we have used a preosteoblastic cell line to study the effects of dynamic mechanical strain on beta-catenin signaling. We found that mechanical strain caused a rapid, transient accumulation of active beta-catenin in the cytoplasm and its translocation to the nucleus. This was followed by up-regulation of the Wnt/beta-catenin target genes Wisp1 and Cox2, with peak responses at 4 and 1 h of strain, respectively. The increase of beta-catenin was temporally related to the activation of Akt and subsequent inactivation of GSK3beta, and caveolin-1 was not required for these molecular events. Application of Dkk-1, which disrupts canonical Wnt/LRP5 signaling, did not block strain-induced nuclear translocation of beta-catenin or up-regulation of Wisp1 and Cox2 expression. Conditions that increased basal beta-catenin levels, such as lithium chloride treatment or repression of caveolin-1 expression, were shown to enhance the effects of strain. In summary, mechanical strain activates Akt and inactivates GSK3beta to allow beta-catenin translocation, and Wnt signaling through LRP5 is not required for these strain-mediated responses. Thus, beta-catenin serves as both a modulator and effector of mechanical signals in bone cells.
Figures


References
-
- Rubin, C., and Lanyon, L. (1984) J. Bone Joint Surg. 66A 397-402 - PubMed
-
- Rubin, J., Fan, X., Biskobing, D., Taylor, W., and Rubin, C. (1999) J. Orthop. Res. 17 639-645 - PubMed
-
- Sawakami, K., Robling, A. G., Ai, M., Pitner, N. D., Liu, D., Warden, S. J., Li, J., Maye, P., Rowe, D. W., Duncan, R. L., Warman, M. L., and Turner, C. H. (2006) J. Biol. Chem. 281 23698-23711 - PubMed
-
- Robinson, J. A., Chatterjee-Kishore, M., Yaworsky, P. J., Cullen, D. M., Zhao, W., Li, C., Kharode, Y., Sauter, L., Babij, P., Brown, E. L., Hill, A. A., Akhter, M. P., Johnson, M. L., Recker, R. R., Komm, B. S., and Bex, F. J. (2006) J. Biol. Chem. 281 31720-31728 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
