

Functional studies of multiple thioredoxins from Mycobacterium tuberculosis
- PMID: 18723612
- PMCID: PMC2580692
- DOI: 10.1128/JB.00159-08
Functional studies of multiple thioredoxins from Mycobacterium tuberculosis
Abstract
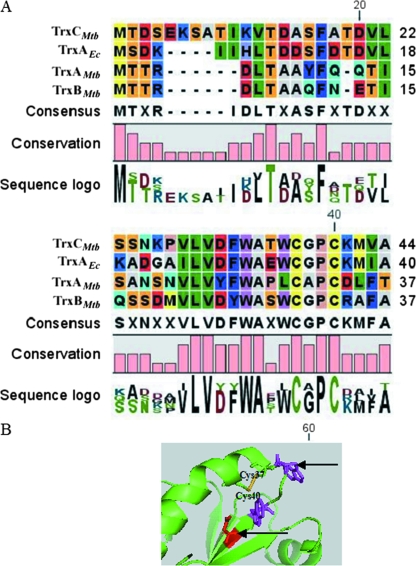
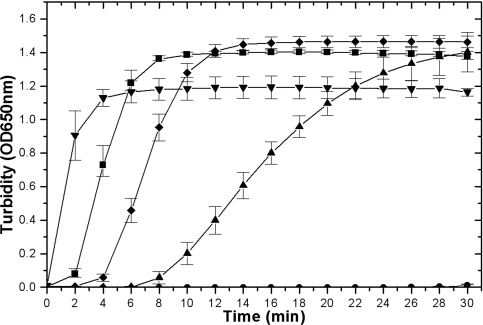
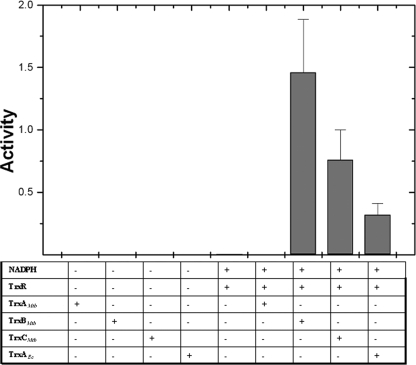
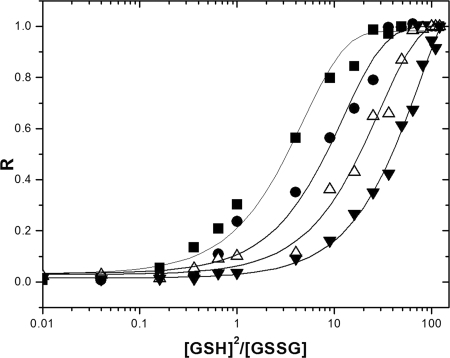
Cytoplasmic protein reduction via generalized thiol/disulfide exchange reactions and maintenance of cellular redox homeostasis is mediated by the thioredoxin superfamily of proteins. Here, we describe the characterization of the thioredoxin system from Mycobacterium tuberculosis, whose genome bears the potential to encode three putative thioredoxins from the open reading frames designated trxAMtb, trxBMtb, and trxCMtb. We show that all three thioredoxins, overproduced in Escherichia coli, are able to reduce insulin, a model substrate, in the presence of dithiothreitol. However, we observe that thioredoxin reductase is not capable of reducing TrxAMtb in an NADPH-dependent manner, indicating that only TrxBMtb and TrxCMtb are the biologically active disulfide reductases. The absence of detectable mRNA transcripts of trxAMtb observed when M. tuberculosis strain H37Rv was cultivated under different growth conditions suggests that trxAMtb expression may be cryptic. The measured redox potentials of TrxBMtb and TrxCMtb (-262+/-2 mV and -269+/-2 mV, respectively) render these proteins somewhat more oxidizing than E. coli thioredoxin 1 (TrxA). In E. coli strains lacking components of cytoplasmic protein reduction pathways, heterologous expression of the mycobacterial thioredoxins was able to effectively substitute for their function.
Figures

Similar articles
-
Protein disulfide-isomerase is a substrate for thioredoxin reductase and has thioredoxin-like activity.J Biol Chem. 1990 Jun 5;265(16):9114-20. J Biol Chem. 1990. PMID: 2188973
-
Redox biology of Mycobacterium tuberculosis H37Rv: protein-protein interaction between GlgB and WhiB1 involves exchange of thiol-disulfide.BMC Biochem. 2009 Jan 5;10:1. doi: 10.1186/1471-2091-10-1. BMC Biochem. 2009. PMID: 19121228 Free PMC article.
-
The thioredoxin system from Streptomyces coelicolor.J Basic Microbiol. 2006;46(1):47-55. doi: 10.1002/jobm.200510036. J Basic Microbiol. 2006. PMID: 16463318
-
The plant thioredoxin system.Cell Mol Life Sci. 2005 Jan;62(1):24-35. doi: 10.1007/s00018-004-4296-4. Cell Mol Life Sci. 2005. PMID: 15619004 Free PMC article. Review.
-
Thioredoxin and related proteins in procaryotes.FEMS Microbiol Rev. 1988 Dec;4(4):271-97. doi: 10.1111/j.1574-6968.1988.tb02747.x. FEMS Microbiol Rev. 1988. PMID: 3152490 Review.
Cited by
-
Solution NMR Studies of Mycobacterium tuberculosis Proteins for Antibiotic Target Discovery.Molecules. 2017 Aug 31;22(9):1447. doi: 10.3390/molecules22091447. Molecules. 2017. PMID: 28858250 Free PMC article. Review.
-
Thioredoxins, glutaredoxins, and peroxiredoxins--molecular mechanisms and health significance: from cofactors to antioxidants to redox signaling.Antioxid Redox Signal. 2013 Nov 1;19(13):1539-605. doi: 10.1089/ars.2012.4599. Epub 2013 Mar 28. Antioxid Redox Signal. 2013. PMID: 23397885 Free PMC article. Review.
-
Ebselen: A Promising Repurposing Drug to Treat Infections Caused by Multidrug-Resistant Microorganisms.Interdiscip Perspect Infect Dis. 2024 Mar 30;2024:9109041. doi: 10.1155/2024/9109041. eCollection 2024. Interdiscip Perspect Infect Dis. 2024. PMID: 38586592 Free PMC article. Review.
-
Mycobacterium tuberculosis Proteome Response to Antituberculosis Compounds Reveals Metabolic "Escape" Pathways That Prolong Bacterial Survival.Antimicrob Agents Chemother. 2017 Jun 27;61(7):e00430-17. doi: 10.1128/AAC.00430-17. Print 2017 Jul. Antimicrob Agents Chemother. 2017. PMID: 28416555 Free PMC article.
-
Functional analysis of thioredoxin from the desert lichen-forming fungus, Endocarpon pusillum Hedwig, reveals its role in stress tolerance.Sci Rep. 2016 Jun 2;6:27184. doi: 10.1038/srep27184. Sci Rep. 2016. PMID: 27251605 Free PMC article.
References
-
- Akif, M., K. Suhre, C. Verma, and S. C. Mande. 2005. Conformational flexibility of Mycobacterium tuberculosis thioredoxin reductase: crystal structure and normal-mode analysis. Acta Crystallogr. D 611603-1611. - PubMed
-
- Arner, E. S. J., and A. Holmgren. 2000. Physiological functions of thioredoxin and thioredoxin reductase. Eur. J. Biochem. 2676102-6109. - PubMed
-
- Åslund, F., K. D. Berndt, and A. Holmgren. 1997. Redox potentials of glutaredoxins and other thiol-disulfide oxidoreductases of the thioredoxin superfamily determined by direct protein-protein redox equilibria. J. Biol. Chem. 27230780-30786. - PubMed
-
- Bardwell, J. C. A., K. McGovern, and J. Beckwith. 1991. Identification of a protein required for disulfide bond formation in-vivo. Cell 67581-589. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
