

A domino effect in antifolate drug action in Escherichia coli
- PMID: 18724364
- PMCID: PMC2699209
- DOI: 10.1038/nchembio.108
A domino effect in antifolate drug action in Escherichia coli
Abstract
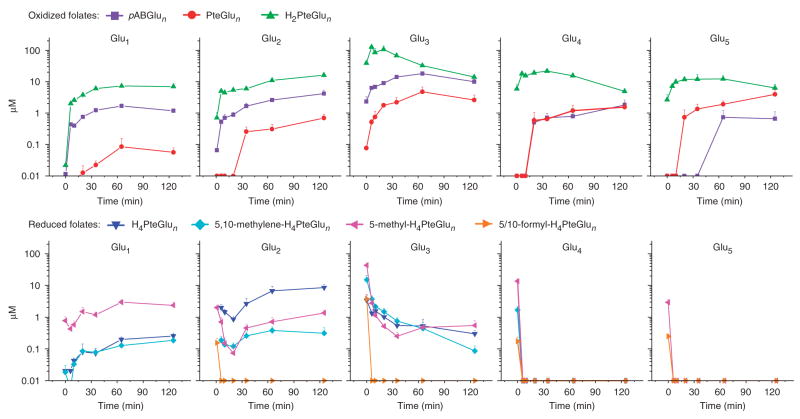
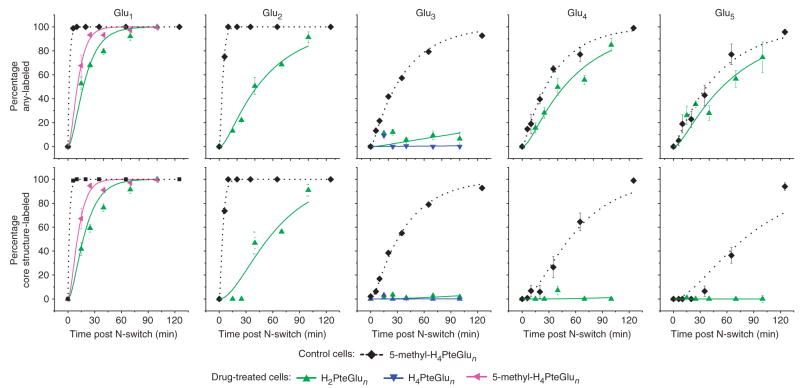
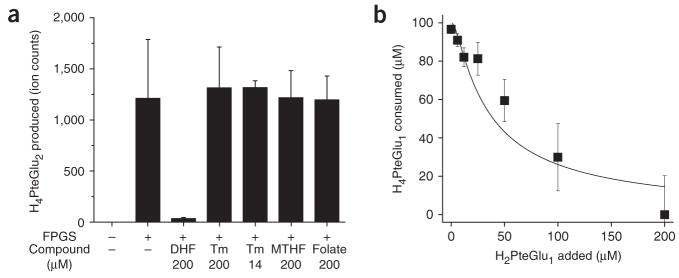
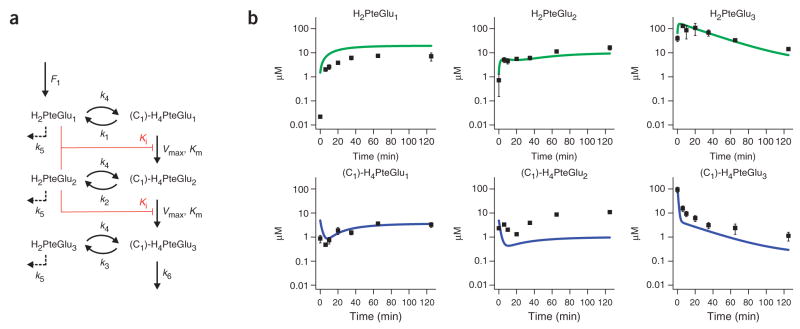
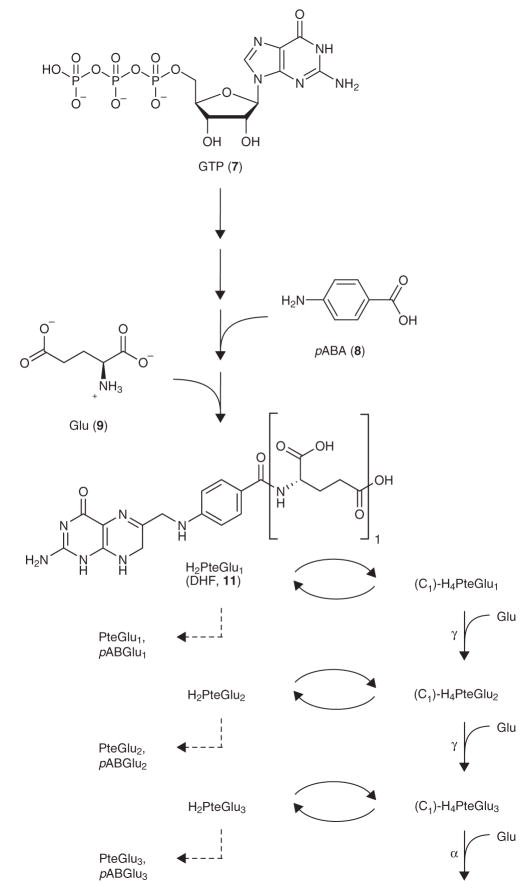
Mass spectrometry technologies for measurement of cellular metabolism are opening new avenues to explore drug activity. Trimethoprim is an antibiotic that inhibits bacterial dihydrofolate reductase (DHFR). Kinetic flux profiling with (15)N-labeled ammonia in Escherichia coli reveals that trimethoprim leads to blockade not only of DHFR but also of another critical enzyme of folate metabolism: folylpoly-gamma-glutamate synthetase (FP-gamma-GS). Inhibition of FP-gamma-GS is not directly due to trimethoprim. Instead, it arises from accumulation of DHFR's substrate dihydrofolate, which we show is a potent FP-gamma-GS inhibitor. Thus, owing to the inherent connectivity of the metabolic network, falling DHFR activity leads to falling FP-gamma-GS activity in a domino-like cascade. This cascade results in complex folate dynamics, and its incorporation in a computational model of folate metabolism recapitulates the dynamics observed experimentally. These results highlight the potential for quantitative analysis of cellular metabolism to reveal mechanisms of drug action.
Figures
References
-
- Voet D, Voet JG. In: Biochemistry. Harris D, Fitzgerald P, editors. John Wiley & Sons, Inc; Hoboken, New Jersey, USA: 2004. pp. 482–486.
-
- Quinlivan EP, McPartlin J, Weir DG, Scott J. Mechanism of the antimicrobial drug trimethoprim revisited. FASEB J. 2000;14:2519–2524. - PubMed
-
- Sabatine MS, et al. Metabolomic identification of novel biomarkers of myocardial ischemia. Circulation. 2005;112:3868–3875. - PubMed
-
- Bajad SU, et al. Separation and quantitation of water soluble cellular metabolites by hydrophilic interaction chromatography-tandem mass spectrometry. J Chromatogr A. 2006;1125:76–88. - PubMed
-
- van der Werf MJ, Overkamp KM, Muilwijk B, Coulier L, Hankemeier T. Microbial metabolomics: toward a platform with full metabolome coverage. Anal Biochem. 2007;370:17–25. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Chemical Information
Molecular Biology Databases
