

Stimulation of Toll-like receptor 3 and 4 induces interleukin-1beta maturation by caspase-8
- PMID: 18725521
- PMCID: PMC2526192
- DOI: 10.1084/jem.20071632
Stimulation of Toll-like receptor 3 and 4 induces interleukin-1beta maturation by caspase-8
Abstract
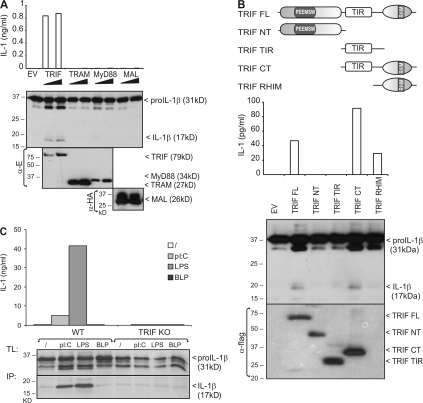
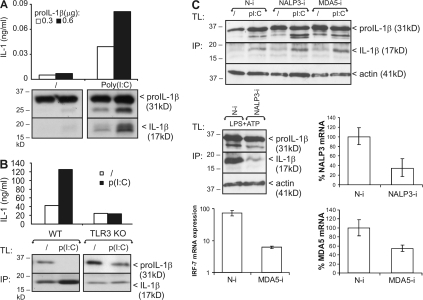
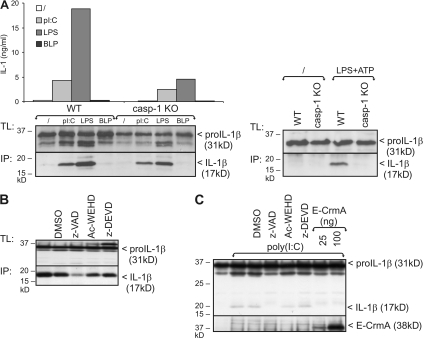
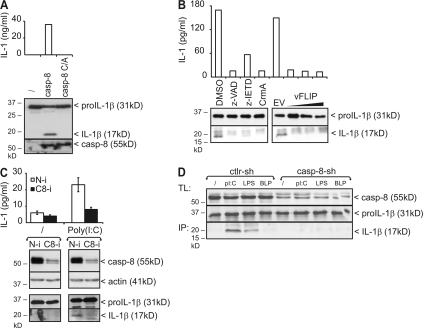
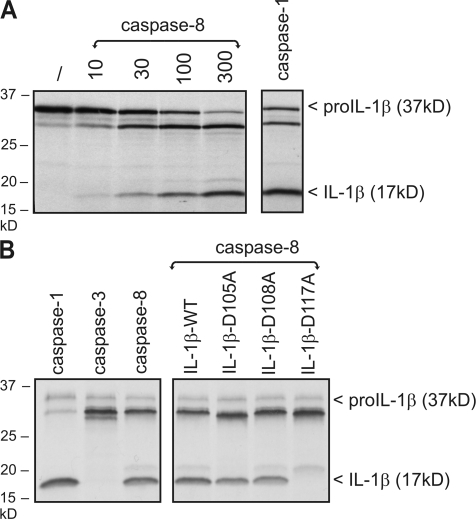
The cytokine interleukin (IL)-1beta is a key mediator of the inflammatory response and has been implicated in the pathophysiology of acute and chronic inflammation. IL-1beta is synthesized in response to many stimuli as an inactive pro-IL-1beta precursor protein that is further processed by caspase-1 into mature IL-1beta, which is the secreted biologically active form of the cytokine. Although stimulation of membrane-bound Toll-like receptors (TLRs) up-regulates pro-IL-1beta expression, activation of caspase-1 is believed to be mainly initiated by cytosolic Nod-like receptors. In this study, we show that polyinosinic:polycytidylic acid (poly[I:C]) and lipopolysaccharide stimulation of macrophages induces pro-IL-1beta processing via a Toll/IL-1R domain-containing adaptor-inducing interferon-beta-dependent signaling pathway that is initiated by TLR3 and TLR4, respectively. Ribonucleic acid interference (RNAi)-mediated knockdown of the intracellular receptors NALP3 or MDA5 did not affect poly(I:C)-induced pro-IL-1beta processing. Surprisingly, poly(I:C)- and LPS-induced pro-IL-1beta processing still occurred in caspase-1-deficient cells. In contrast, pro-IL-1beta processing was inhibited by caspase-8 peptide inhibitors, CrmA or vFLIP expression, and caspase-8 knockdown via RNAi, indicating an essential role for caspase-8. Moreover, recombinant caspase-8 was able to cleave pro-IL-1beta in vitro at exactly the same site as caspase-1. These results implicate a novel role for caspase-8 in the production of biologically active IL-1beta in response to TLR3 and TLR4 stimulation.
Figures
References
-
- Dunne, A., and L.A. O'Neill. 2003. The interleukin-1 receptor/Toll-like receptor superfamily: signal transduction during inflammation and host defense. Sci. STKE. 2003:re3. - PubMed
-
- Martinon, F., and J. Tschopp. 2007. Inflammatory caspases and inflammasomes: master switches of inflammation. Cell Death Differ. 14:10–22. - PubMed
-
- Lamkanfi, M., N. Festjens, W. Declercq, T. Vanden Berghe, and P. Vandenabeele. 2007. Caspases in cell survival, proliferation and differentiation. Cell Death Differ. 14:44–55. - PubMed
-
- Creagh, E.M., and L.A. O'Neill. 2006. TLRs, NLRs and RLRs: a trinity of pathogen sensors that co-operate in innate immunity. Trends Immunol. 27:352–357. - PubMed
-
- Kanneganti, T.D., M. Lamkanfi, Y.G. Kim, G. Chen, J.H. Park, L. Franchi, P. Vandenabeele, and G. Nunez. 2007. Pannexin-1-mediated recognition of bacterial molecules activates the cryopyrin inflammasome independent of Toll-like receptor signaling. Immunity. 26:433–443. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
