

Micropuncturing the nephron
- PMID: 18752000
- PMCID: PMC2954491
- DOI: 10.1007/s00424-008-0581-7
Micropuncturing the nephron
Abstract
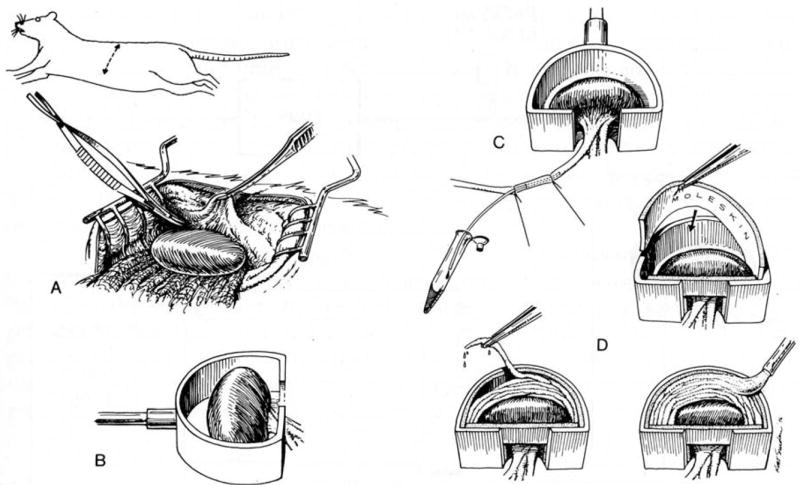
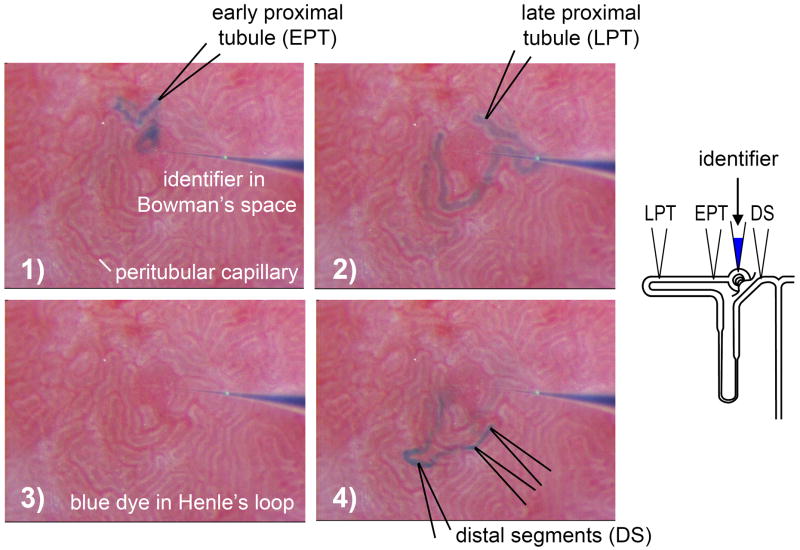
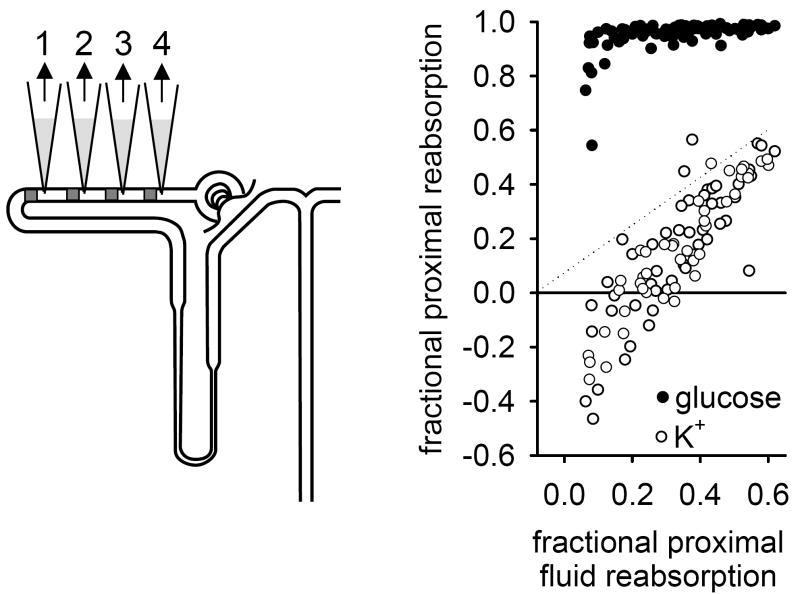
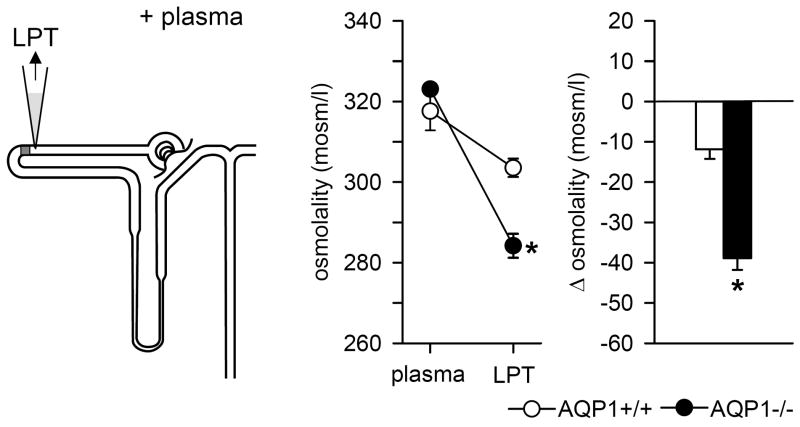
To achieve the role of the kidney in maintaining body homeostasis, the renal vasculature, the glomeruli, and the various segments of the nephron and the collecting duct system have to fulfill very diverse and specific functions. These functions are dependent on a complex renal architecture and are regulated by systemic hemodynamics, hormones, and nerves. As a consequence, to better understand the physiology of the kidney, methods are necessary that allow insights on the function of these diverse structures in the physiological context of the intact kidney. The renal micropuncture technique allows direct access to study superficial nephrons in vivo. In this review, the application of micropuncture techniques on the single nephron level is outlined as an approach to better understand aspects of glomerular filtration, tubular transport, and tubulo-glomerular communication. Studies from the author's lab, including experiments in gene-targeted mice, are briefly presented to illustrate some of the approaches and show how they can further advance our understanding of the molecular mechanisms involved in the regulation of kidney function.
Figures


Similar articles
-
Micropuncture of the kidney: a primer on techniques.Compr Physiol. 2012 Jan;2(1):621-37. doi: 10.1002/cphy.c110035. Compr Physiol. 2012. PMID: 23728982 Review.
-
The influence of tubulo-glomerular feedback on the autoregulation of filtration rate in superficial and deep glomeruli.Acta Physiol Scand. 1984 Nov;122(3):235-42. doi: 10.1111/j.1748-1716.1984.tb07506.x. Acta Physiol Scand. 1984. PMID: 6516878
-
The tubulo-glomerular feedback mechanism-a determinant for the autoregulation of the glomerular filtration rate in superficial and juxtamedullary nephrons.Klin Wochenschr. 1982 Sep 15;60(18):1071-6. doi: 10.1007/BF01715837. Klin Wochenschr. 1982. PMID: 7144054
-
Renal potassium homeostasis: a short historical perspective.Semin Nephrol. 2013 May;33(3):209-14. doi: 10.1016/j.semnephrol.2013.05.001. Semin Nephrol. 2013. PMID: 23953798
-
Renal potassium transport: contributions of individual nephron segments and populations.Am J Physiol. 1978 Dec;235(6):F515-27. doi: 10.1152/ajprenal.1978.235.6.F515. Am J Physiol. 1978. PMID: 367178 Review.
Cited by
-
Mapping single-nephron filtration in the isolated, perfused rat kidney using magnetic resonance imaging.Am J Physiol Renal Physiol. 2022 Nov 1;323(5):F602-F611. doi: 10.1152/ajprenal.00103.2022. Epub 2022 Sep 1. Am J Physiol Renal Physiol. 2022. PMID: 36049066 Free PMC article.
-
Net K+ secretion in the thick ascending limb of mice on a low-Na, high-K diet.Kidney Int. 2017 Oct;92(4):864-875. doi: 10.1016/j.kint.2017.04.009. Epub 2017 Jul 6. Kidney Int. 2017. PMID: 28688582 Free PMC article.
-
Proximal nephron.Compr Physiol. 2013 Jul;3(3):1079-123. doi: 10.1002/cphy.c110061. Compr Physiol. 2013. PMID: 23897681 Free PMC article. Review.
-
Proteomic analysis of murine kidney proximal tubule sub-segment derived cell lines reveals preferences in mitochondrial pathway activity.J Proteomics. 2023 Oct 30;289:104998. doi: 10.1016/j.jprot.2023.104998. Epub 2023 Aug 30. J Proteomics. 2023. PMID: 37657718 Free PMC article.
-
Cross-Species Insights into Autosomal Dominant Polycystic Kidney Disease: Provide an Alternative View on Research Advancement.Int J Mol Sci. 2024 May 22;25(11):5646. doi: 10.3390/ijms25115646. Int J Mol Sci. 2024. PMID: 38891834 Free PMC article. Review.
References
-
- Andreucci VE, Herrera-Acosta J, Rector FC, Jr, Seldin DW. Measurement of single-nephron glomerular filtration rate by micropuncture: analysis of error. Am J Physiol. 1971;221:1551–1559. - PubMed
-
- Ashworth SL, Sandoval RM, Tanner GA, Molitoris BA. Two-photon microscopy: visualization of kidney dynamics. Kidney Int. 2007;72:416–421. - PubMed
-
- Bailey MA, Unwin RJ, Shirley DG. In vivo inhibition of renal 11beta-hydroxysteroid dehydrogenase in the rat stimulates collecting duct sodium reabsorption. Clin Sci (Lond) 2001;101:195–198. - PubMed
-
- Blantz RC, Tucker BJ. Measurements of glomerular dynamics. In: Martinez-Maldonado, editor. Methods in pharmacology. Plenum Publishing Corporation; 1978. pp. 141–163.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
