

Developmental excitation of corticothalamic neurons by nicotinic acetylcholine receptors
- PMID: 18753377
- PMCID: PMC2909269
- DOI: 10.1523/JNEUROSCI.2645-08.2008
Developmental excitation of corticothalamic neurons by nicotinic acetylcholine receptors
Abstract
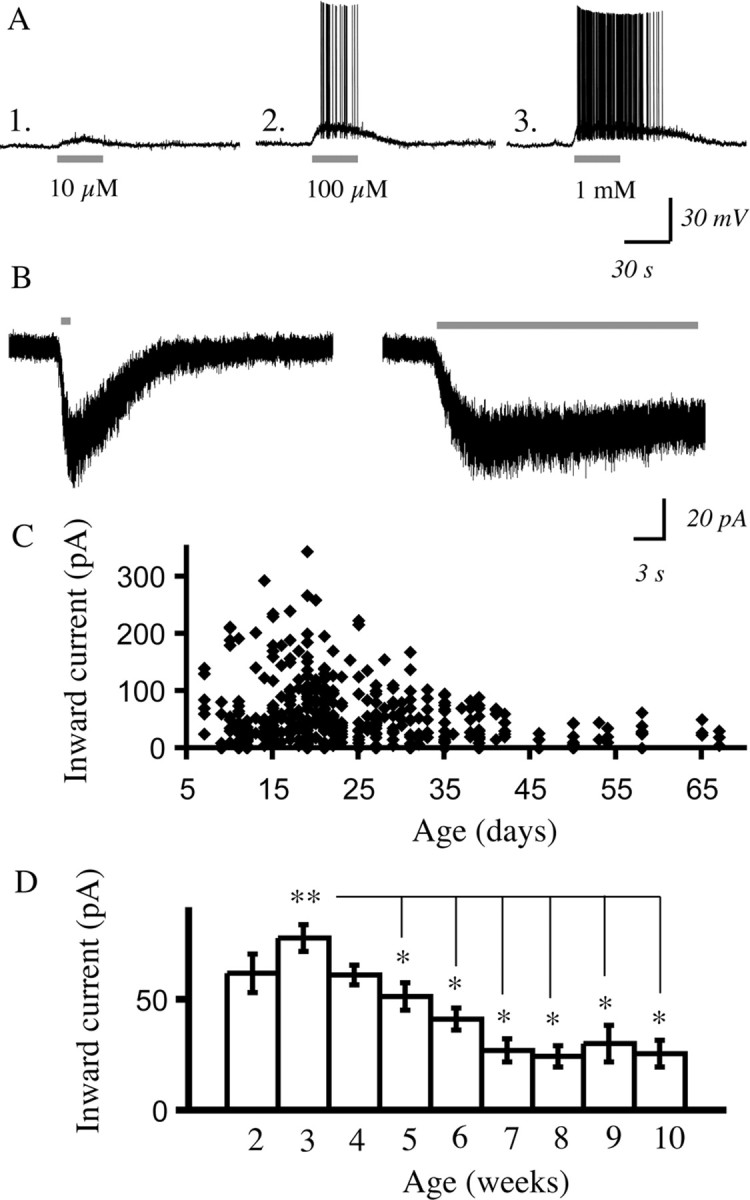
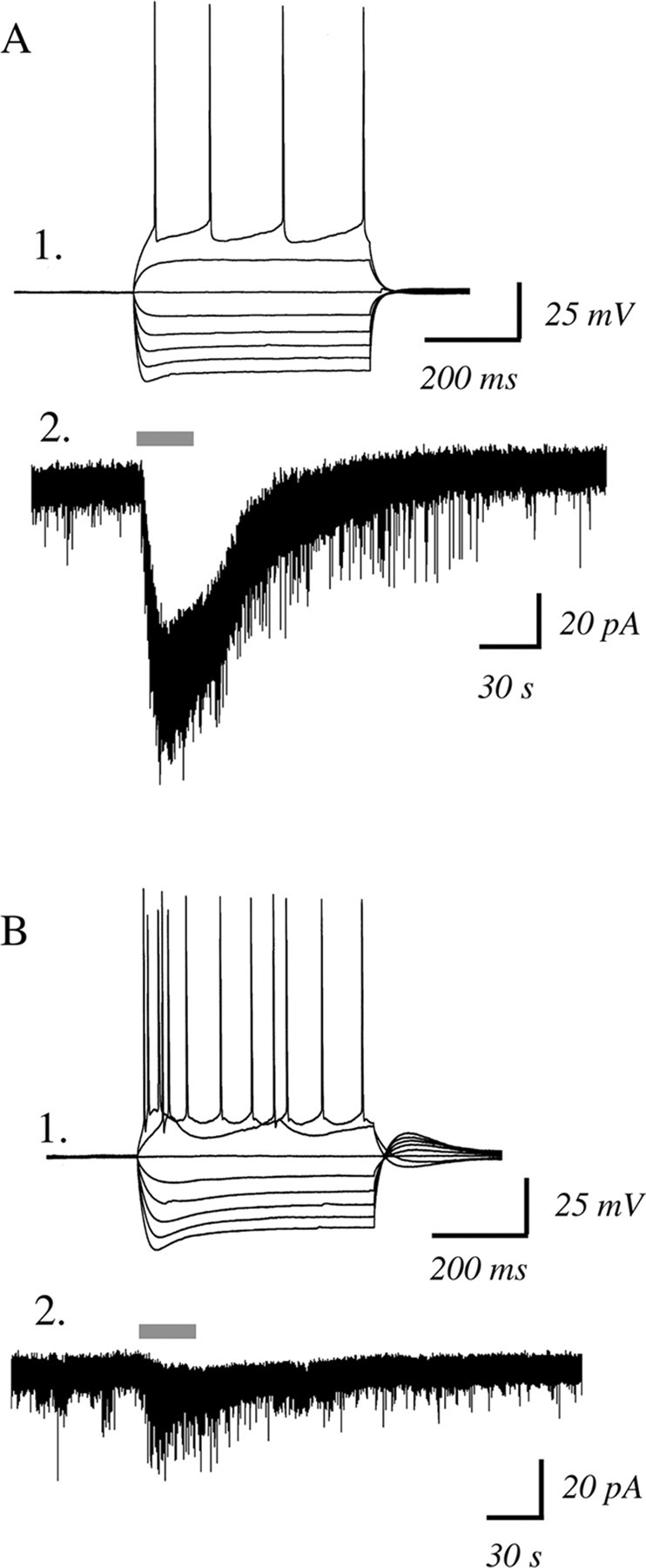
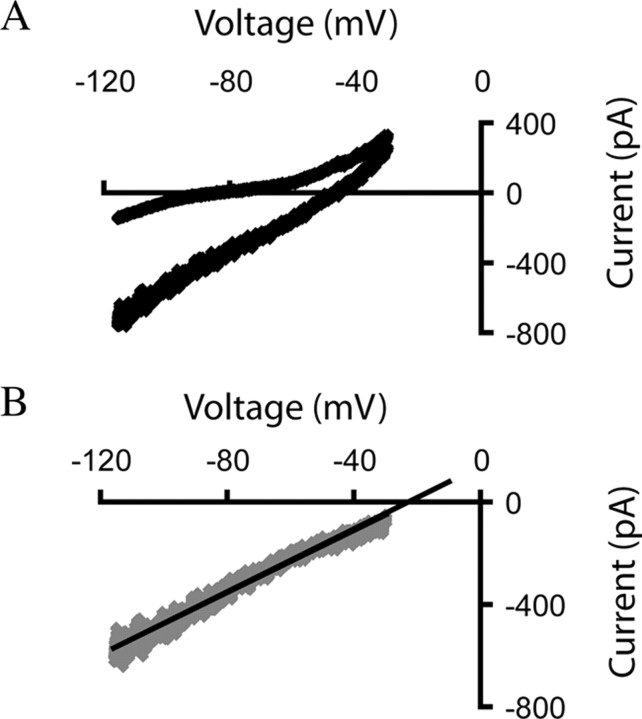
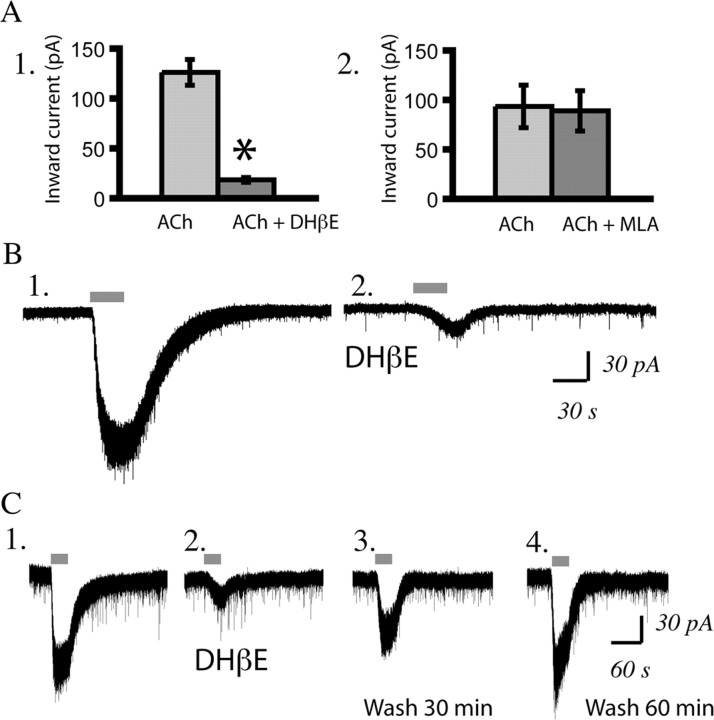
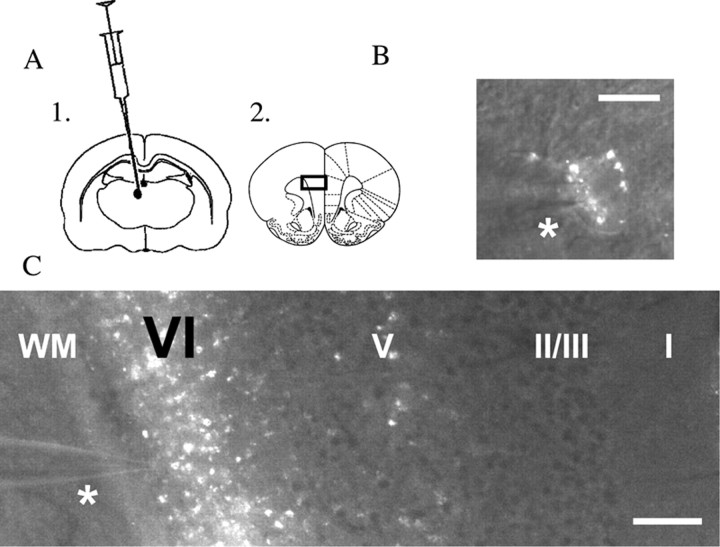
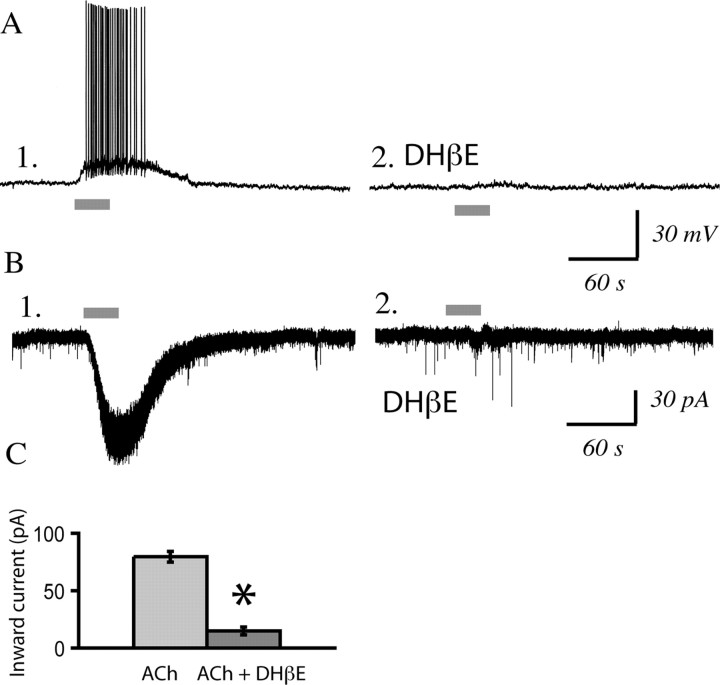
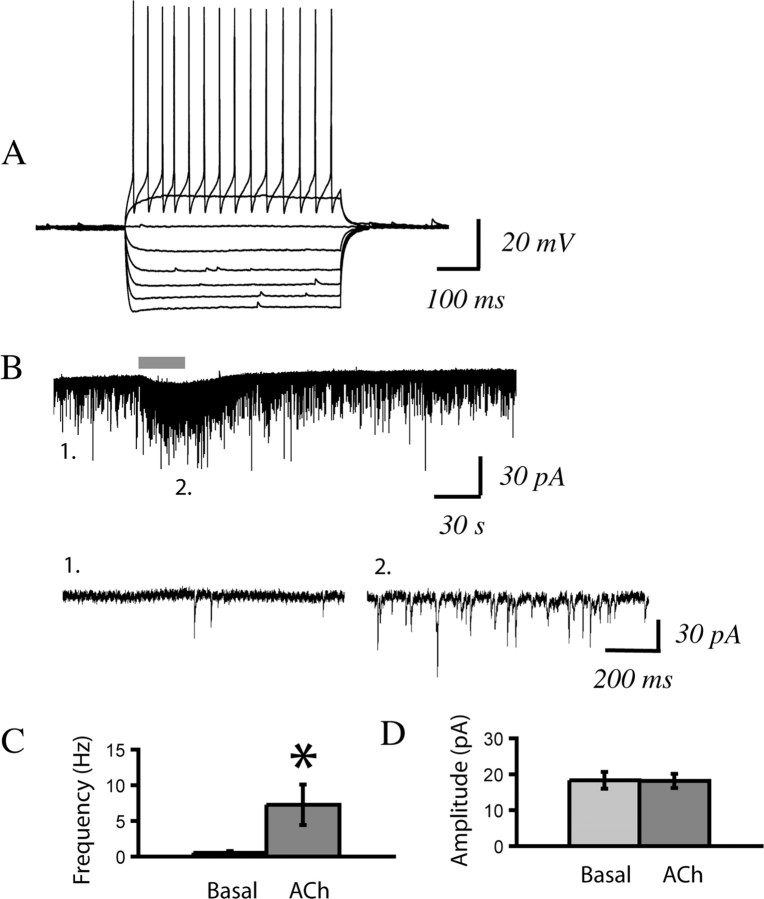
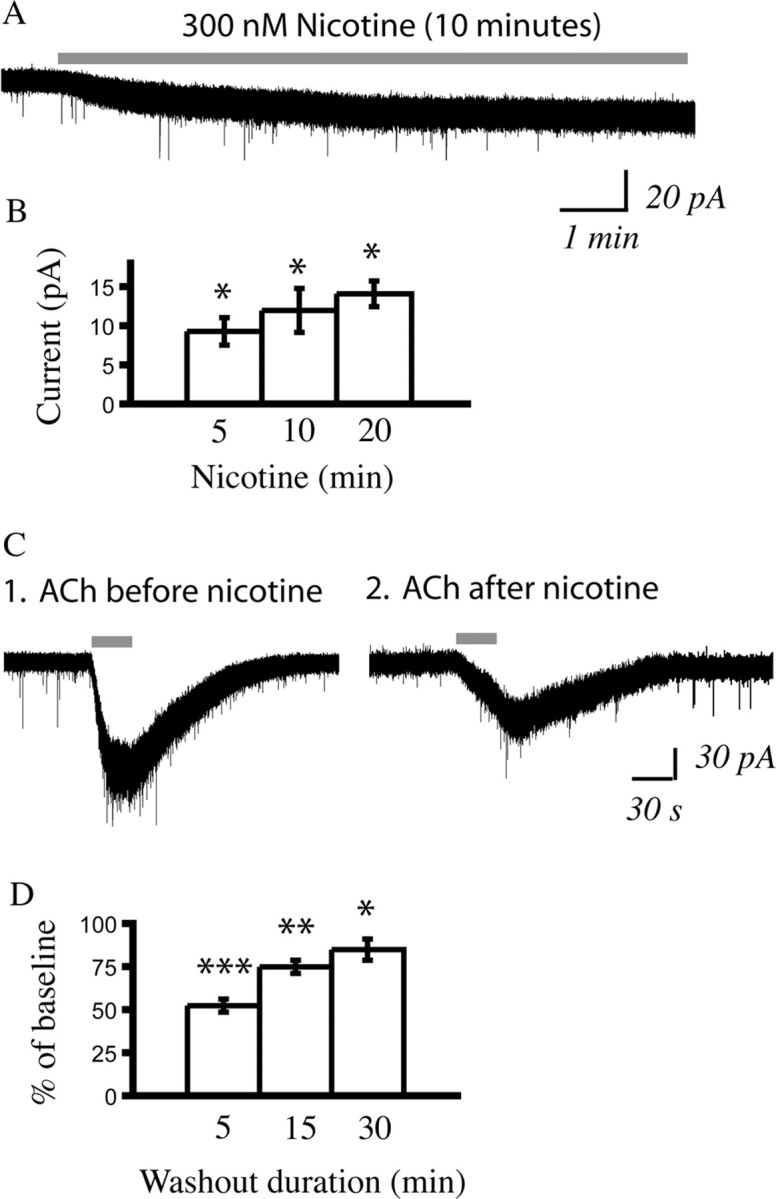
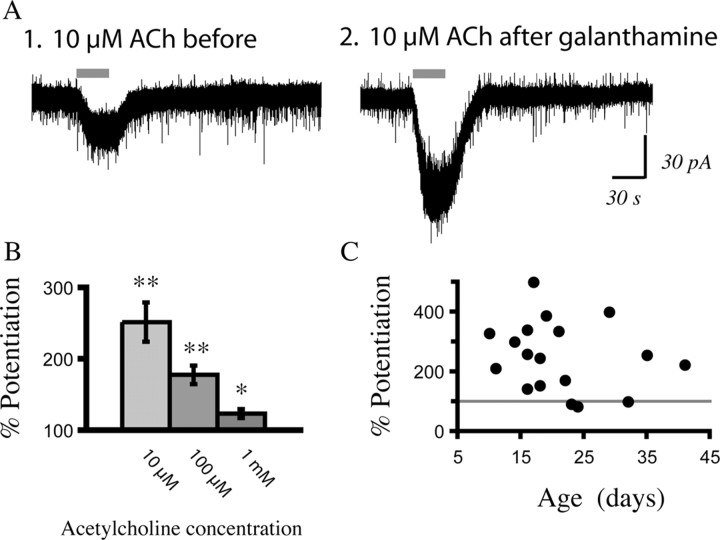
In this study, we show robust nicotinic excitation of pyramidal neurons in layer VI of prefrontal cortex. This layer contains the corticothalamic neurons, which gate thalamic activity and play a critical role in attention. Our experiments tested nicotinic excitation across postnatal development, using whole-cell recordings in prefrontal brain slices from rats. These experiments showed that layer VI neurons have peak nicotinic currents during the first postnatal month, a time period of intensive cortical development in rodents. We demonstrate that these currents are mediated directly by postsynaptic nicotinic receptors and can be suppressed by a competitive antagonist of alpha(4)beta(2)* nicotinic receptors. To record from identified corticothalamic neurons, we performed stereotaxic surgery to label the neurons projecting to medial dorsal thalamus. As hypothesized, recordings from these retrogradely labeled neurons in layer VI showed prominent nicotinic currents. Finally, we examined the effects of the drug nicotine on layer VI neurons and probed for the potential involvement of the accessory subunit, alpha(5), in their receptors. A level of nicotine similar to that found in the blood of smokers elicits a stable inward current in layer VI neurons, yet this exposure desensitizes approximately 50% of the subsequent current elicited by acetylcholine. An allosteric modulator of alpha(4)beta(2)alpha(5) receptors resulted in a 2.5-fold potentiation of submaximal nicotinic currents. This result is consistent with the expression of the relatively rare alpha(5) nicotinic subunit in layer VI. In summary, we show that layer VI corticothalamic neurons can be strongly excited during development by an unusual subtype of nicotinic receptor.
Figures
References
-
- Belluardo N, Mudò G, Blum M, Cheng Q, Caniglia G, Dell'Albani P, Fuxe K. The nicotinic acetylcholine receptor agonist (+/−)-epibatidine increases FGF-2 mRNA and protein levels in the rat brain. Brain Res Mol Brain Res. 1999;74:98–110. - PubMed
-
- Bierut LJ, Stitzel JA, Wang JC, Hinrichs AL, Grucza RA, Xuei X, Saccone NL, Saccone SF, Bertelsen S, Fox L, Horton WJ, Breslau N, Budde J, Cloninger CR, Dick DM, Foroud T, Hatsukami D, Hesselbrock V, Johnson EO, Kramer J, Kuperman S, Madden PA, Mayo K, Nurnberger J, Jr, Pomerleau O, Porjesz B, Reyes O, Schuckit M, Swan G, Tischfield JA, Edenberg HJ, Rice JP, Goate AM. Variants in nicotinic receptors and risk for nicotine dependence. Am J Psychiatry. 2008 in press. - PMC - PubMed
-
- Breese CR, Lee MJ, Adams CE, Sullivan B, Logel J, Gillen KM, Marks MJ, Collins AC, Leonard S. Abnormal regulation of high affinity nicotinic receptors in subjects with schizophrenia. Neuropsychopharmacology. 2000;23:351–364. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources