

Evolution rescues folding of human immunodeficiency virus-1 envelope glycoprotein GP120 lacking a conserved disulfide bond
- PMID: 18753405
- PMCID: PMC2575144
- DOI: 10.1091/mbc.e08-07-0670
Evolution rescues folding of human immunodeficiency virus-1 envelope glycoprotein GP120 lacking a conserved disulfide bond
Abstract
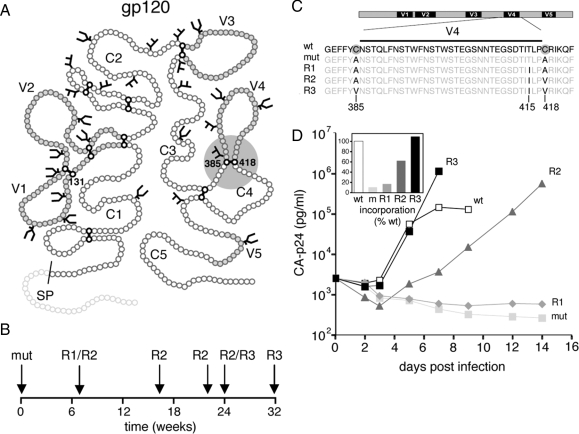
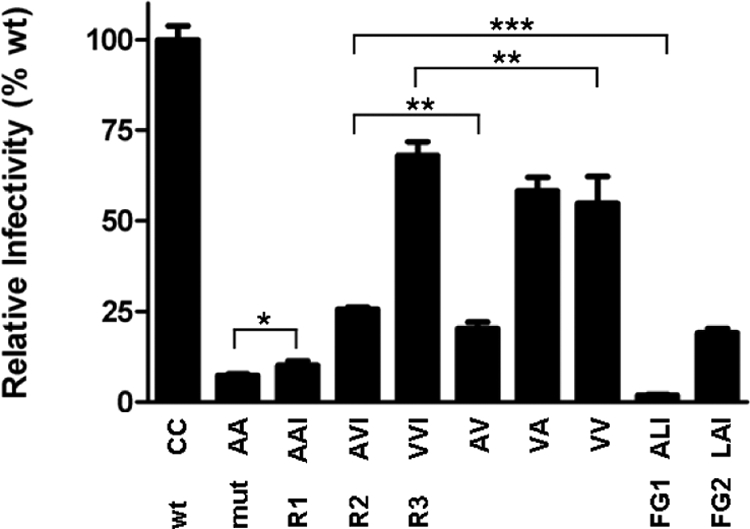
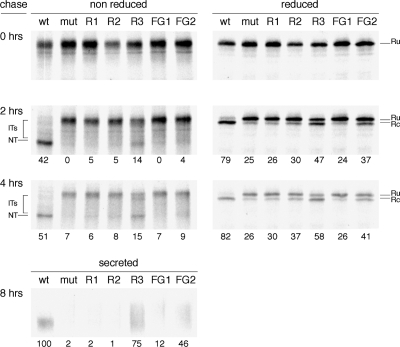
The majority of eukaryotic secretory and membrane proteins contain disulfide bonds, which are strongly conserved within protein families because of their crucial role in folding or function. The exact role of these disulfide bonds during folding is unclear. Using virus-driven evolution we generated a viral glycoprotein variant, which is functional despite the lack of an absolutely conserved disulfide bond that links two antiparallel beta-strands in a six-stranded beta-barrel. Molecular dynamics simulations revealed that improved hydrogen bonding and side chain packing led to stabilization of the beta-barrel fold, implying that beta-sheet preference codirects glycoprotein folding in vivo. Our results show that the interactions between two beta-strands that are important for the formation and/or integrity of the beta-barrel can be supported by either a disulfide bond or beta-sheet favoring residues.
Figures
References
-
- Berendsen H.J.C., Postma J.P.M., Van Gunsteren W. F., Di-Nola A., Haak J. R. Molecular dynamics with coupling to an external bath. J. Chem. Phys. 1984;81:3684–3690.
-
- Berendsen H J.C., Postma J.P.M., Van Gunsteren W. F., Hermans J. Interaction Models for Water in Relation to Protein Hydration. Dordrecht, The Netherlands: Reidel Publishing; 1981.
-
- Chen B., Vogan E. M., Gong H., Skehel J. J., Wiley D. C., Harrison S. C. Structure of an unliganded simian immunodeficiency virus gp120 core. Nature. 2005;433:834–841. - PubMed
-
- Chou P. Y., Fasman G. D. Conformational parameters for amino acids in helical, beta-sheet, and random coil regions calculated from proteins. Biochemistry. 1974;13:211–222. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
