

Epileptiform activities in slices of hippocampus from mice after intra-hippocampal injection of kainic acid
- PMID: 18755752
- PMCID: PMC2614071
- DOI: 10.1113/jphysiol.2008.156281
Epileptiform activities in slices of hippocampus from mice after intra-hippocampal injection of kainic acid
Abstract
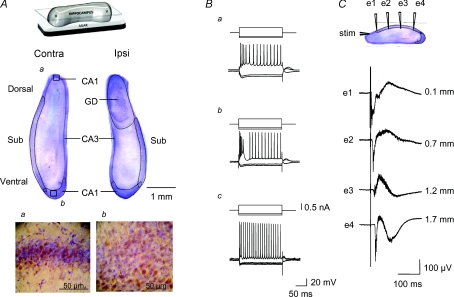
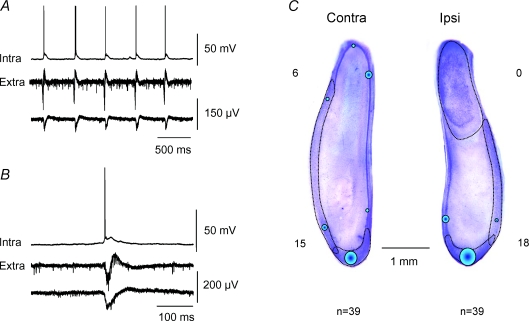
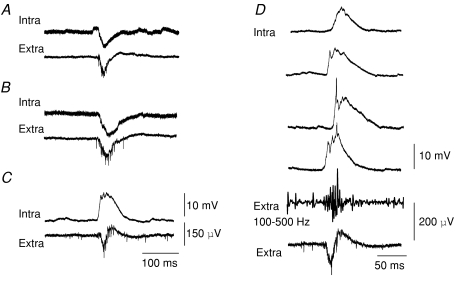
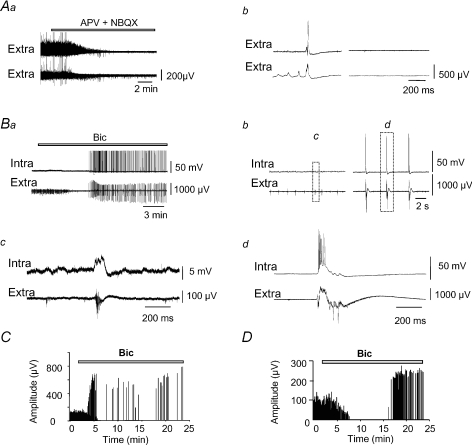
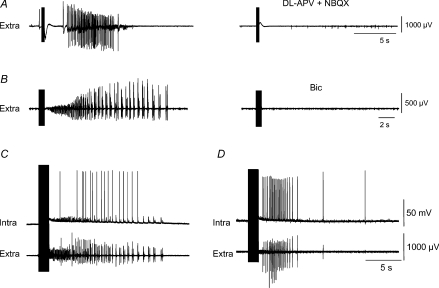
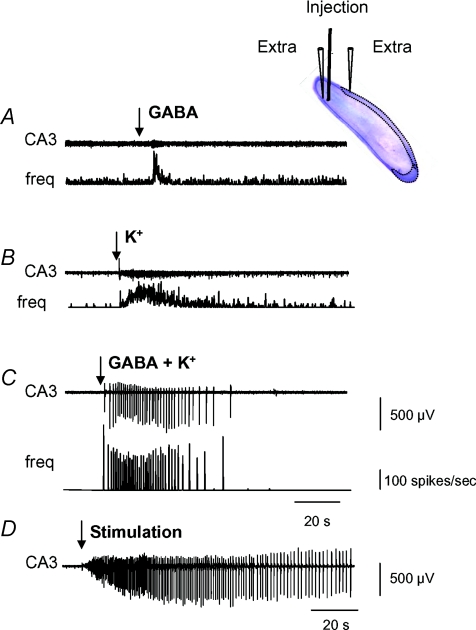
Intra-hippocampal kainate injection induces the emergence of recurrent seizures after a delay of 3-4 weeks. We examined the cellular and synaptic basis of this activity in vitro using extracellular and intracellular records from longitudinal hippocampal slices. These slices permitted recordings from the dentate gyrus, the CA3 and CA1 regions and the subiculum of both the injected and the contralateral non-injected hippocampus. A sclerotic zone was evident in dorsal regions of slices from the injected hippocampus, while ventral regions and tissue from the contralateral hippocampus were not sclerotic. Interictal field potentials of duration 50-200 ms were generated spontaneously in both ipsilateral and contralateral hippocampal slices, but not in the sclerotic region, at 3-12 months after injection. They were initiated in the CA1 and CA3 regions and the subiculum. They were blocked by antagonists at glutamatergic receptors and were transformed into prolonged epileptiform events by GABAergic receptor antagonists. The membrane potential and the reversal potential of GABAergic synaptic events were more depolarized in CA1 pyramidal cells from kainate-treated animals than in control animals. Ictal-like events of duration 8-80 s were induced by tetanic stimulation (50 Hz, 0.2-1 s) preferentially in dorsal contralateral and ventral ipsilateral slices. Similar events were initiated by focal application of a combination of high K(+) and GABA. These data show that both interictal and ictal-like activities can be induced in slices of both ipsilateral and contralateral hippocampus from kainate-treated animals and suggest that changes in cellular excitability and inhibitory synaptic signalling may contribute to their generation.
Figures


Similar articles
-
Effects of focal injection of kainic acid into the mouse hippocampus in vitro and ex vivo.J Physiol. 2005 Dec 15;569(Pt 3):833-47. doi: 10.1113/jphysiol.2005.094599. Epub 2005 Oct 20. J Physiol. 2005. PMID: 16239280 Free PMC article.
-
Ictal epileptiform activity in the CA3 region of hippocampal slices produced by pilocarpine.J Neurophysiol. 1998 Jun;79(6):3019-29. doi: 10.1152/jn.1998.79.6.3019. J Neurophysiol. 1998. PMID: 9636105
-
Local circuit abnormalities in chronically epileptic rats after intrahippocampal tetanus toxin injection in infancy.J Neurophysiol. 1998 Jan;79(1):106-16. doi: 10.1152/jn.1998.79.1.106. J Neurophysiol. 1998. PMID: 9425181
-
Interictal-ictal interactions and limbic seizure generation.Rev Neurol (Paris). 1999 Jul;155(6-7):468-71. Rev Neurol (Paris). 1999. PMID: 10472661 Review.
-
Network and pharmacological mechanisms leading to epileptiform synchronization in the limbic system in vitro.Prog Neurobiol. 2002 Oct;68(3):167-207. doi: 10.1016/s0301-0082(02)00077-1. Prog Neurobiol. 2002. PMID: 12450487 Review.
Cited by
-
Progressive, potassium-sensitive epileptiform activity in hippocampal area CA3 of pilocarpine-treated rats with recurrent seizures.Epilepsy Res. 2011 Nov;97(1-2):92-102. doi: 10.1016/j.eplepsyres.2011.07.008. Epub 2011 Aug 30. Epilepsy Res. 2011. PMID: 21880468 Free PMC article.
-
Increased Dentate Gyrus Excitability in the Intrahippocampal Kainic Acid Mouse Model for Temporal Lobe Epilepsy.Int J Mol Sci. 2024 Jan 4;25(1):660. doi: 10.3390/ijms25010660. Int J Mol Sci. 2024. PMID: 38203829 Free PMC article.
-
Disrupted Co-activation of Interneurons and Hippocampal Network after Focal Kainate Lesion.Front Neural Circuits. 2017 Nov 13;11:87. doi: 10.3389/fncir.2017.00087. eCollection 2017. Front Neural Circuits. 2017. PMID: 29180954 Free PMC article.
-
Involvement of the thalamic parafascicular nucleus in mesial temporal lobe epilepsy.J Neurosci. 2010 Dec 8;30(49):16523-35. doi: 10.1523/JNEUROSCI.1109-10.2010. J Neurosci. 2010. PMID: 21147992 Free PMC article.
-
Immediate Epileptogenesis after Kainate-Induced Status Epilepticus in C57BL/6J Mice: Evidence from Long Term Continuous Video-EEG Telemetry.PLoS One. 2015 Jul 10;10(7):e0131705. doi: 10.1371/journal.pone.0131705. eCollection 2015. PLoS One. 2015. PMID: 26161754 Free PMC article.
References
-
- Andersson PB, Perry VH, Gordon S. The CNS acute inflammatory response to excitotoxic neuronal cell death. Immunol Lett. 1991;30:177–181. - PubMed
-
- Becker AJ, Chen J, Zien A, Sochivko D, Normann S, Schramm J, Elger CE, Wiestler OD, Blumcke I. Correlated stage- and subfield-associated hippocampal gene expression patterns in experimental and human temporal lobe epilepsy. Eur J Neurosci. 2003;18:2792–2802. - PubMed
-
- Ben-Ari Y. Limbic seizure and brain damage produced by kainic acid: mechanisms and relevance to human temporal lobe epilepsy. Neuroscience. 1985;14:375–403. - PubMed
-
- Ben-Ari Y, Cossart R. Kainate, a double agent that generates seizures: two decades of progress. Trends Neurosci. 2000;23:580–587. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous