

Modulation of Runx2 activity by estrogen receptor-alpha: implications for osteoporosis and breast cancer
- PMID: 18755791
- PMCID: PMC2613062
- DOI: 10.1210/en.2008-0680
Modulation of Runx2 activity by estrogen receptor-alpha: implications for osteoporosis and breast cancer
Abstract
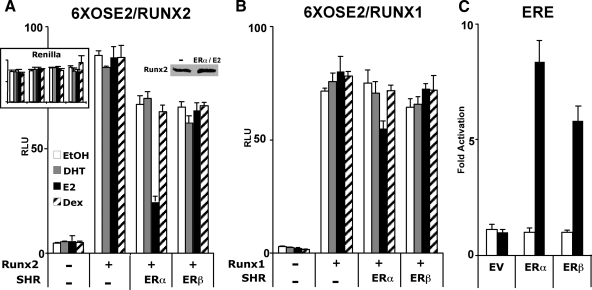
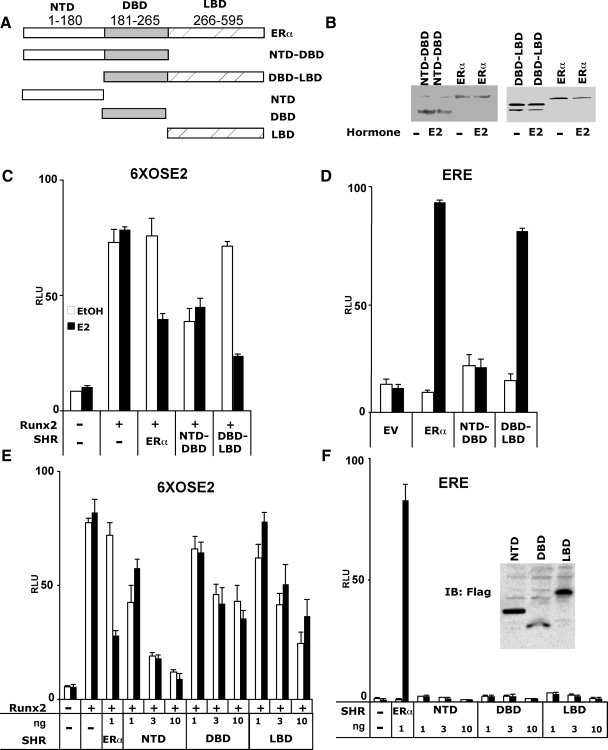
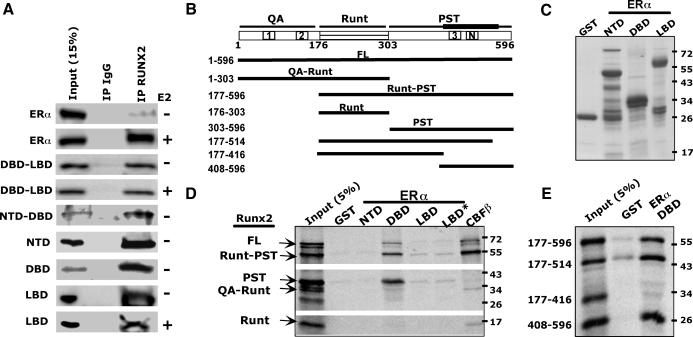
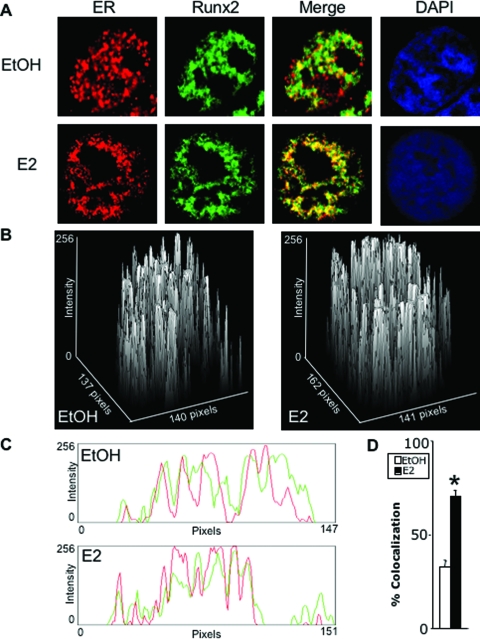
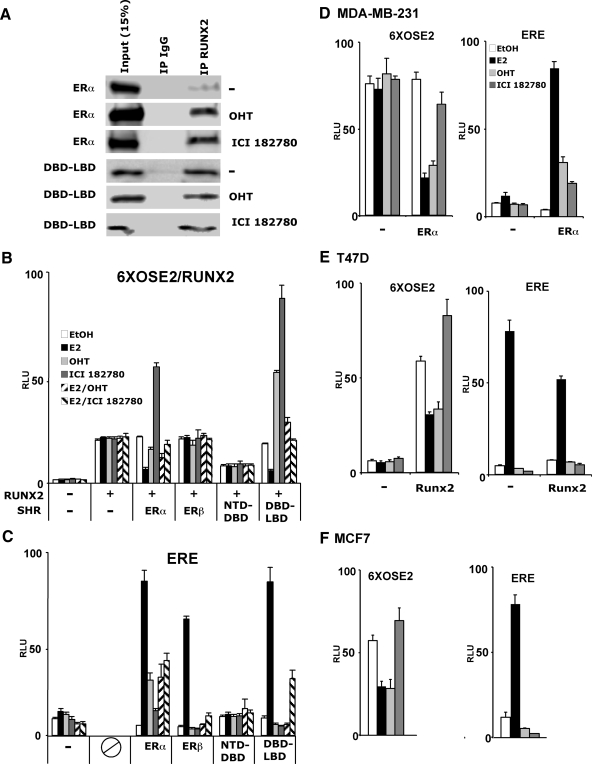
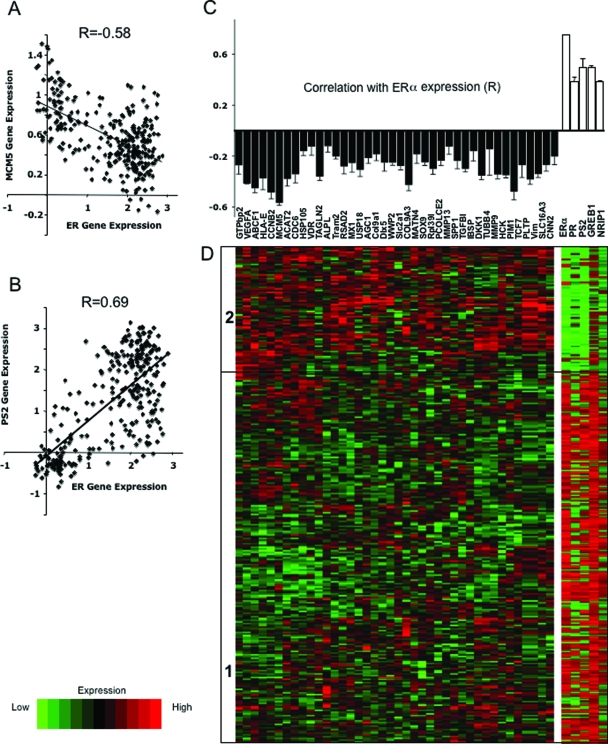
The transcription factors Runx2 and estrogen receptor-alpha (ERalpha) are involved in numerous normal and disease processes, including postmenopausal osteoporosis and breast cancer. Using indirect immunofluorescence microscopy and pull-down techniques, we found them to colocalize and form complexes in a ligand-dependent manner. Estradiol-bound ERalpha strongly interacted with Runx2 directly through its DNA-binding domain and only indirectly through its N-terminal and ligand-binding domains. Runx2's amino acids 417-514, encompassing activation domain 3 and the nuclear matrix targeting sequence, were sufficient for interaction with ERalpha's DNA-binding domain. As a consequence of the interaction, Runx2's transcriptional activation activity was strongly repressed, as shown by reporter assays in COS7 cells, breast cancer cells, and late-stage MC3T3-E1 osteoblast cultures. Metaanalysis of gene expression in 779 breast cancer biopsies indicated negative correlation between the expression of ERalpha and Runx2 target genes. Selective ER modulators (SERM) induced ERalpha-Runx2 interactions but led to various functional outcomes. The regulation of Runx2 by ERalpha may play key roles in osteoblast and breast epithelial cell growth and differentiation; hence, modulation of Runx2 by native and synthetic ERalpha ligands offers new avenues in selective ER modulator evaluation and development.
Figures

References
-
- Coffman JA 2003 Runx transcription factors and the developmental balance between cell proliferation and differentiation. Cell Biol Int 27:315–324 - PubMed
-
- Schroeder TM, Jensen ED, Westendorf JJ 2005 Runx2: a master organizer of gene transcription in developing and maturing osteoblasts. Birth Defects Res C Embryo Today 75:213–225 - PubMed
-
- Komori T, Yagi H, Nomura S, Yamaguchi A, Sasaki K, Deguchi K, Shimizu Y, Bronson RT, Gao YH, Inada M, Sato M, Okamoto R, Kitamura Y, Yoshiki S, Kishimoto T 1997 Targeted disruption of Cbfa1 results in a complete lack of bone formation owing to maturational arrest of osteoblasts. Cell 89:755–764 - PubMed
-
- Otto F, Thornell AP, Crompton T, Denzel A, Gilmour KC, Rosewell IR, Stamp GW, Beddington RS, Mundlos S, Olsen BR, Selby PB, Owen MJ 1997 Cbfa1, a candidate gene for cleidocranial dysplasia syndrome, is essential for osteoblast differentiation and bone development. Cell 89:765–771 - PubMed
-
- Salingcarnboriboon R, Tsuji K, Komori T, Nakashima K, Ezura Y, Noda M 2006 Runx2 is a target of mechanical unloading to alter osteoblastic activity and bone formation in vivo. Endocrinology 147:2296–2305 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
