

EBV latent membrane protein 1 effects on plakoglobin, cell growth, and migration
- PMID: 18757414
- PMCID: PMC2593097
- DOI: 10.1158/0008-5472.CAN-08-1178
EBV latent membrane protein 1 effects on plakoglobin, cell growth, and migration
Abstract
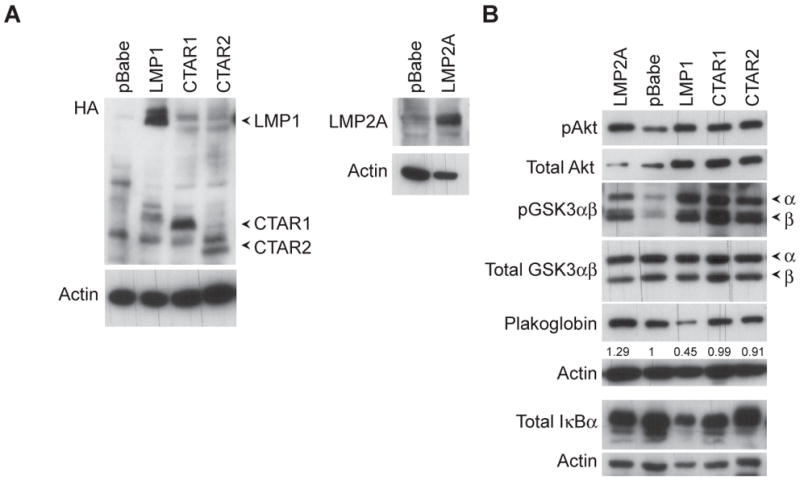
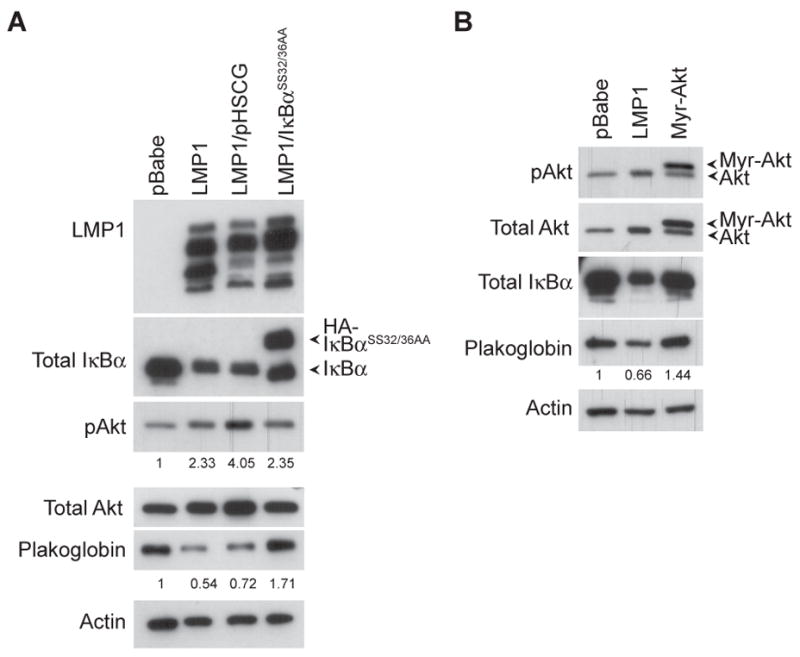
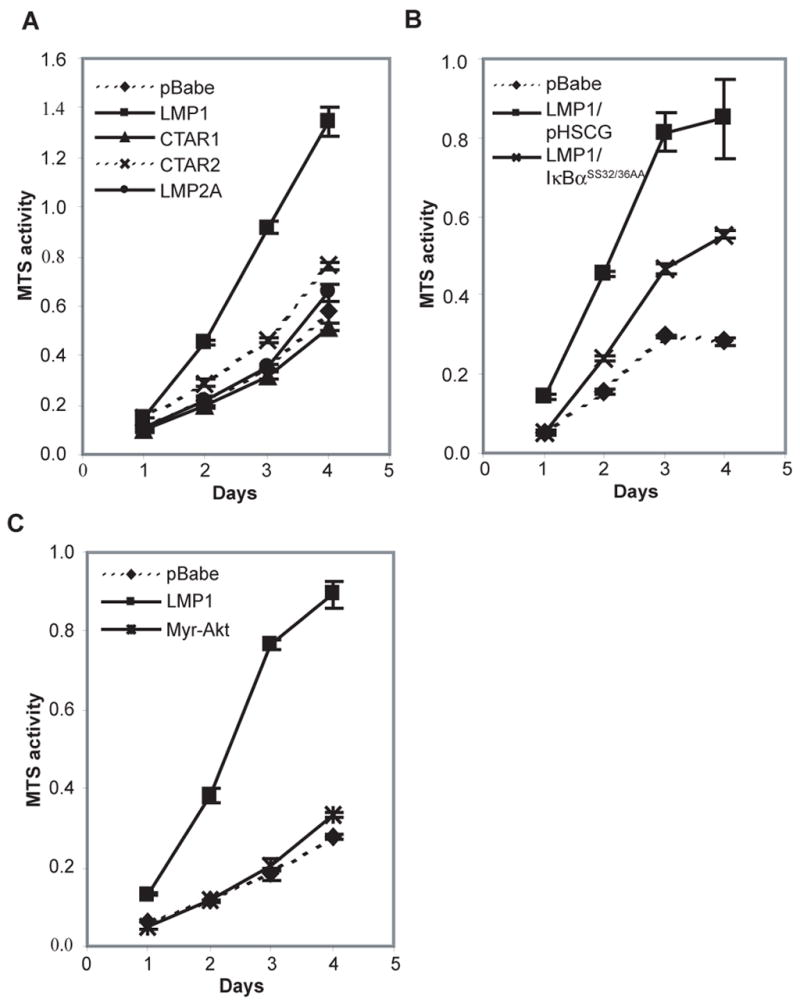
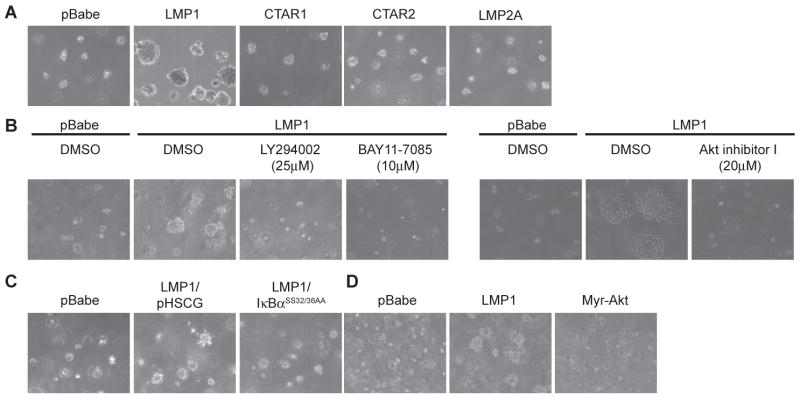
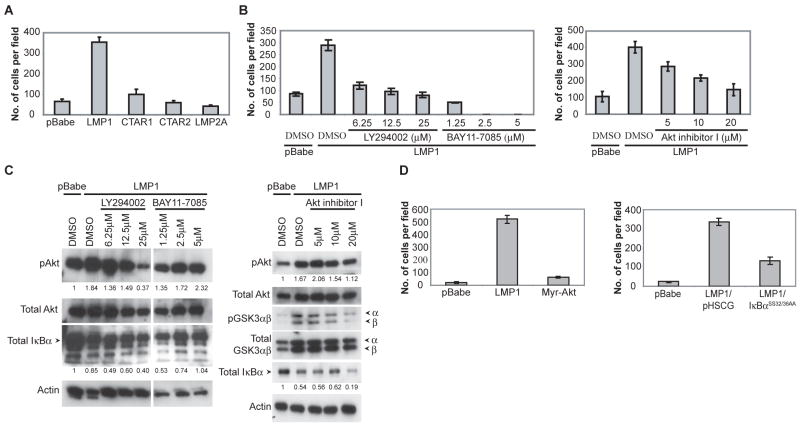
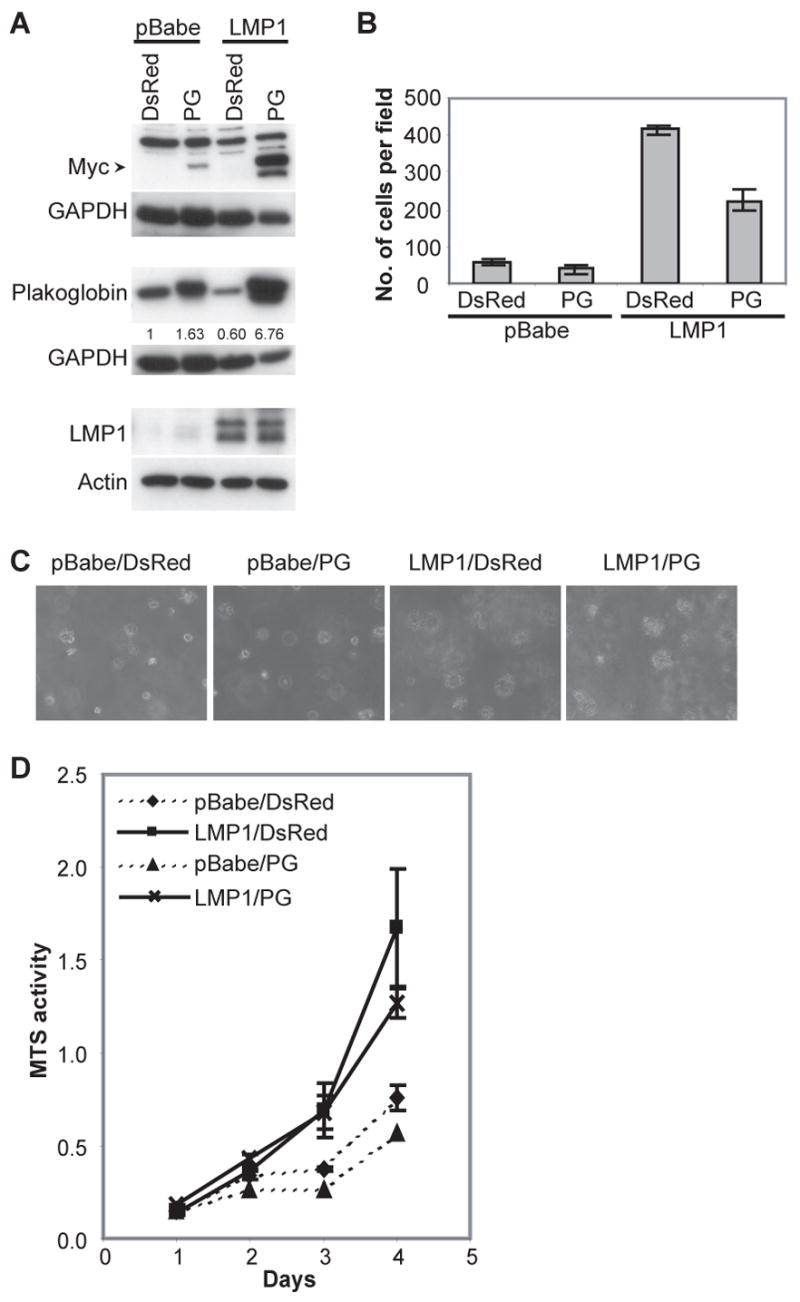
Latent membrane protein 1 (LMP1), the major oncoprotein of EBV, is likely responsible for many of the altered cellular growth properties in EBV-associated cancers, including nasopharyngeal carcinoma (NPC). In this study, the effects of LMP1 on cell growth and migration were studied in the context of the EBV-positive C666-1 NPC cell line. In the soft agar transformation and Transwell metastasis assays, LMP1 enhanced cell growth and migration through activation of phosphatidylinositol 3-kinase (PI3K)/Akt and nuclear factor-kappaB (NF-kappaB) signaling. Inhibitors of PI3K, Akt, and NF-kappaB signaling dramatically reduced these enhanced properties. An IkappaBalpha super-repressor also blocked these effects. However, constitutive activation of Akt alone did not alter cell growth, suggesting that both PI3K/Akt and NF-kappaB activation are required by LMP1. These enhanced effects required the full-length LMP1 encompassing both the PI3K/Akt-activating COOH-terminal activation region (CTAR) 1 and the nonredundant NF-kappaB-activating regions CTAR1 and CTAR2. LMP2A, a latent protein that is also frequently expressed in NPC, similarly activates the PI3K/Akt pathway; however, its overexpression in C666-1 cells did not affect cell growth or migration. LMP1 also decreased expression of the junctional protein plakoglobin, which was shown to be partially responsible for enhanced migration induced by LMP1. This study reveals that in epithelial cells the transforming properties of LMP1 require activation of both PI3K/Akt and NF-kappaB and shows that the loss of plakoglobin expression by LMP1 is a significant factor in the enhanced migration.
Figures
Similar articles
-
Epstein-Barr virus latent membrane protein-1 effects on junctional plakoglobin and induction of a cadherin switch.Cancer Res. 2009 Jul 15;69(14):5734-42. doi: 10.1158/0008-5472.CAN-09-0468. Epub 2009 Jul 7. Cancer Res. 2009. PMID: 19584275 Free PMC article.
-
Unique signaling properties of CTAR1 in LMP1-mediated transformation.J Virol. 2007 Sep;81(18):9680-92. doi: 10.1128/JVI.01001-07. Epub 2007 Jul 11. J Virol. 2007. PMID: 17626074 Free PMC article.
-
Epstein-Barr virus latent membrane protein 1 CTAR1 mediates rodent and human fibroblast transformation through activation of PI3K.Oncogene. 2005 Oct 20;24(46):6917-24. doi: 10.1038/sj.onc.1208846. Oncogene. 2005. PMID: 16007144
-
LMP1 TRAFficking activates growth and survival pathways.Adv Exp Med Biol. 2007;597:173-87. doi: 10.1007/978-0-387-70630-6_14. Adv Exp Med Biol. 2007. PMID: 17633026 Review.
-
The significance of LMP1 expression in nasopharyngeal carcinoma.Semin Cancer Biol. 2002 Dec;12(6):473-87. doi: 10.1016/s1044579x02000901. Semin Cancer Biol. 2002. PMID: 12450733 Review.
Cited by
-
Interaction of phospholipid scramblase 1 with the Epstein-Barr virus protein BZLF1 represses BZLF1-mediated lytic gene transcription.J Biol Chem. 2019 Oct 11;294(41):15104-15116. doi: 10.1074/jbc.RA119.008193. Epub 2019 Aug 21. J Biol Chem. 2019. PMID: 31434743 Free PMC article.
-
The Epstein-Barr virus LMP1 interactome: biological implications and therapeutic targets.Future Virol. 2018 Dec;13(12):863-887. doi: 10.2217/fvl-2018-0120. Epub 2018 Dec 3. Future Virol. 2018. PMID: 34079586 Free PMC article.
-
LMP1-induced cell death may contribute to the emergency of its oncogenic property.PLoS One. 2013 Apr 23;8(4):e60743. doi: 10.1371/journal.pone.0060743. Print 2013. PLoS One. 2013. PMID: 23637765 Free PMC article.
-
EBV Association with Lymphomas and Carcinomas in the Oral Compartment.Viruses. 2022 Dec 1;14(12):2700. doi: 10.3390/v14122700. Viruses. 2022. PMID: 36560704 Free PMC article. Review.
-
LMP1 Up-regulates Calreticulin to Induce Epithelial-mesenchymal Transition via TGF-β/Smad3/NRP1 Pathway in Nasopharyngeal Carcinoma Cells.J Cancer. 2020 Jan 1;11(5):1257-1269. doi: 10.7150/jca.37415. eCollection 2020. J Cancer. 2020. PMID: 31956372 Free PMC article.
References
-
- Raab-Traub N. Pathogenesis of Epstein-Barr virus and its associated malignancies. Semin in virology. 1996;7:315–23.
-
- Rickinson A, Kieff E. Epstein-Barr Virus and Its Replication. In: Knipe ID, editor. Field's Virology. 4. Philadelphia, PA: Lippincott Williams & Wilkins Publishers; 2001. pp. 2511–73.
-
- Mainou BA, Everly DN, Jr, Raab-Traub N. Epstein-Barr virus latent membrane protein 1 CTAR1 mediates rodent and human fibroblast transformation through activation of PI3K. Oncogene. 2005;24:6917–24. - PubMed
-
- Wang D, Liebowitz D, Kieff E. An EBV membrane protein expressed in immortalized lymphocytes transforms established rodent cells. Cell. 1985;43:831–40. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
