

The milkweed pod1 gene encodes a KANADI protein that is required for abaxial/adaxial patterning in maize leaves
- PMID: 18757553
- PMCID: PMC2553616
- DOI: 10.1105/tpc.108.059709
The milkweed pod1 gene encodes a KANADI protein that is required for abaxial/adaxial patterning in maize leaves
Abstract
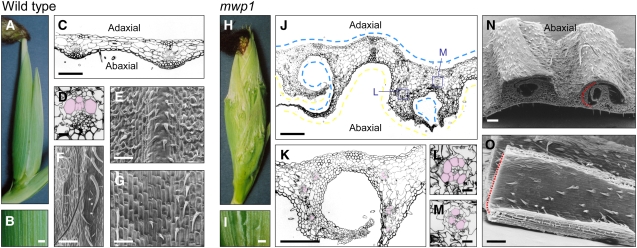
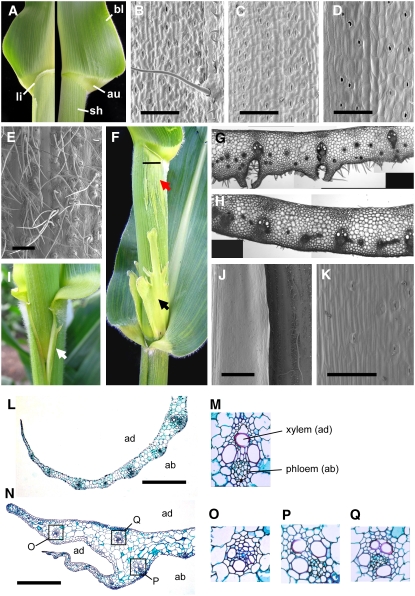
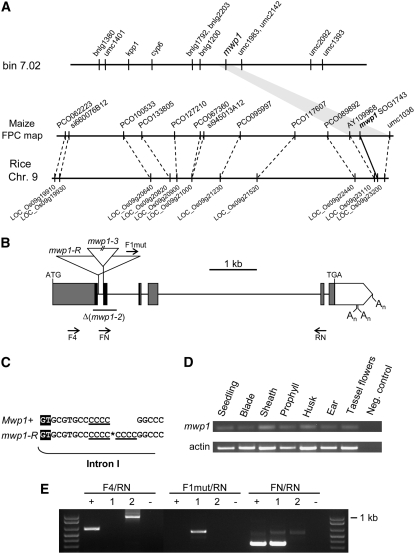
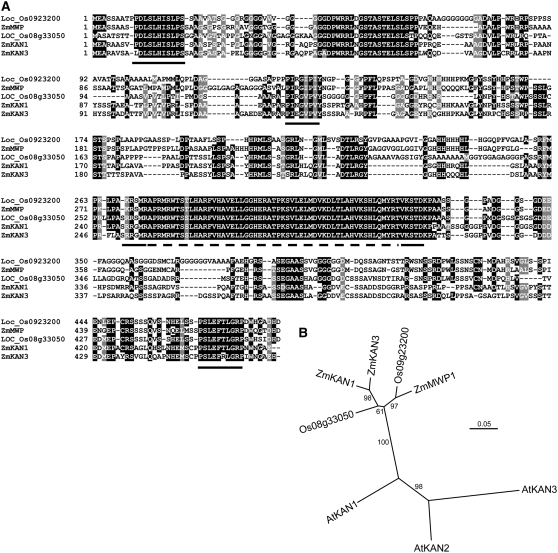
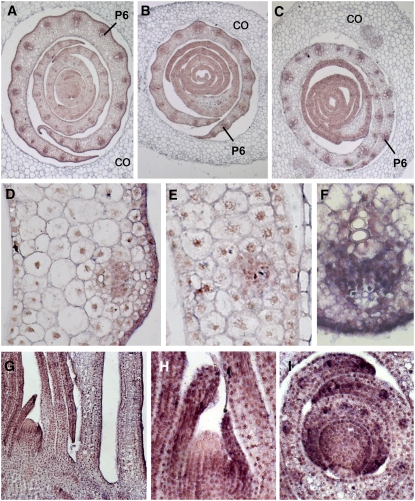
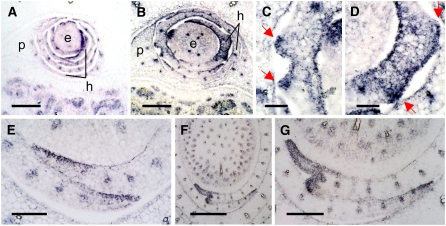
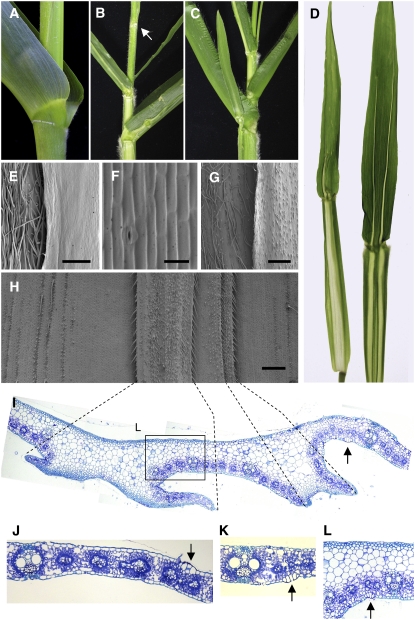
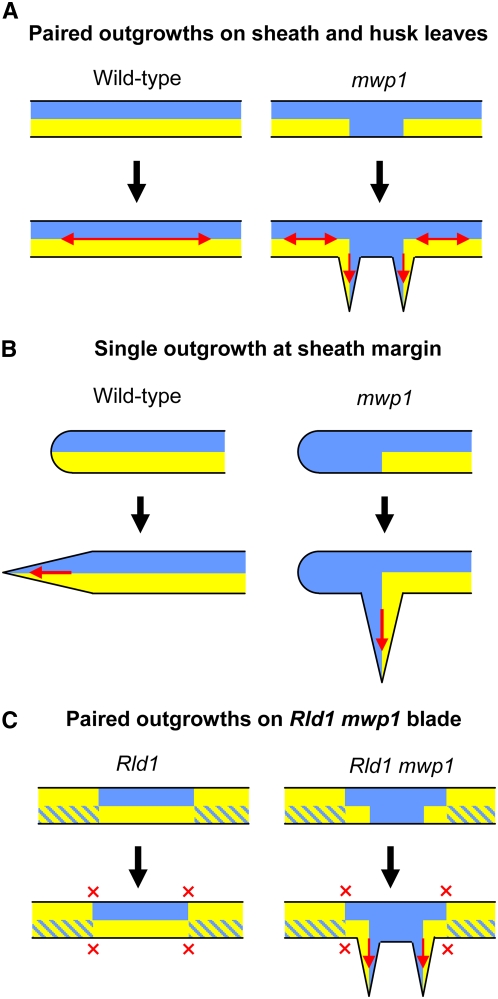
Leaf primordia initiate from the shoot apical meristem with inherent polarity; the adaxial side faces the meristem, while the abaxial side faces away from the meristem. Adaxial/abaxial polarity is thought to be necessary for laminar growth of leaves, as mutants lacking either adaxial or abaxial cell types often develop radially symmetric lateral organs. The milkweed pod1 (mwp1) mutant of maize (Zea mays) has adaxialized sectors in the sheath, the proximal part of the leaf. Ectopic leaf flaps develop where adaxial and abaxial cell types juxtapose. Ectopic expression of the HD-ZIPIII gene rolled leaf1 (rld1) correlates with the adaxialized regions. Cloning of mwp1 showed that it encodes a KANADI transcription factor. Double mutants of mwp1-R with a microRNA-resistant allele of rld1, Rld1-N1990, show a synergistic phenotype with polarity defects in sheath and blade and a failure to differentiate vascular and photosynthetic cell types in the adaxialized sectors. The sectored phenotype and timing of the defect suggest that mwp1 is required late in leaf development to maintain abaxial cell fate. The phenotype of mwp1; Rld1 double mutants shows that both genes are also required early in leaf development to delineate leaf margins as well as to initiate vascular and photosynthetic tissues.
Figures
References
-
- Bao, N., Lye, K.W., and Barton, M.K. (2004). MicroRNA binding sites in Arabidopsis class III HD-ZIP mRNAs are required for methylation of the template chromosome. Dev. Cell 7 653–662. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
