

The crucial role of the Pls1 tetraspanin during ascospore germination in Podospora anserina provides an example of the convergent evolution of morphogenetic processes in fungal plant pathogens and saprobes
- PMID: 18757568
- PMCID: PMC2568061
- DOI: 10.1128/EC.00149-08
The crucial role of the Pls1 tetraspanin during ascospore germination in Podospora anserina provides an example of the convergent evolution of morphogenetic processes in fungal plant pathogens and saprobes
Abstract
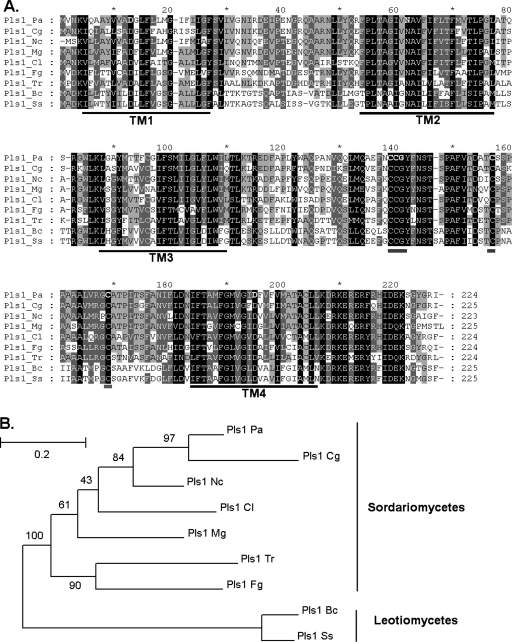
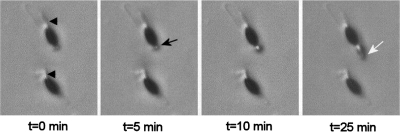
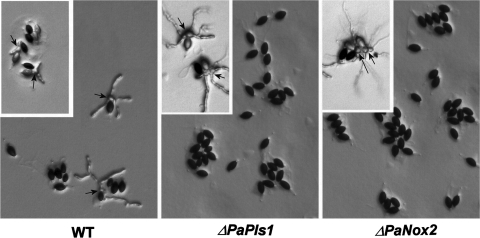
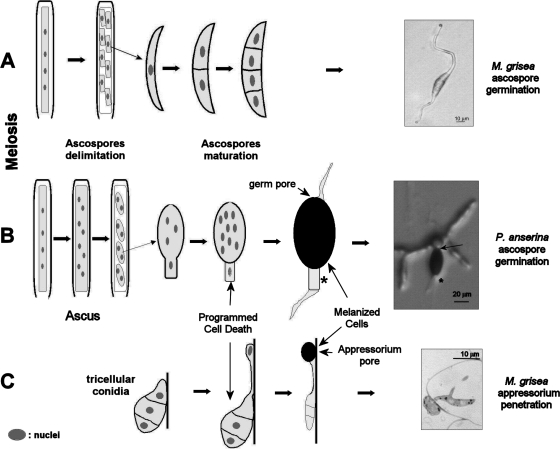
Pls1 tetraspanins were shown for some pathogenic fungi to be essential for appressorium-mediated penetration into their host plants. We show here that Podospora anserina, a saprobic fungus lacking appressorium, contains PaPls1, a gene orthologous to known PLS1 genes. Inactivation of PaPls1 demonstrates that this gene is specifically required for the germination of ascospores in P. anserina. These ascospores are heavily melanized cells that germinate under inducing conditions through a specific pore. On the contrary, MgPLS1, which fully complements a DeltaPaPls1 ascospore germination defect, has no role in the germination of Magnaporthe grisea nonmelanized ascospores but is required for the formation of the penetration peg at the pore of its melanized appressorium. P. anserina mutants with mutation of PaNox2, which encodes the NADPH oxidase of the NOX2 family, display the same ascospore-specific germination defect as the DeltaPaPls1 mutant. Both mutant phenotypes are suppressed by the inhibition of melanin biosynthesis, suggesting that they are involved in the same cellular process required for the germination of P. anserina melanized ascospores. The analysis of the distribution of PLS1 and NOX2 genes in fungal genomes shows that they are either both present or both absent. These results indicate that the germination of P. anserina ascospores and the formation of the M. grisea appressorium penetration peg use the same molecular machinery that includes Pls1 and Nox2. This machinery is specifically required for the emergence of polarized hyphae from reinforced structures such as appressoria and ascospores. Its recurrent recruitment during fungal evolution may account for some of the morphogenetic convergence observed in fungi.
Figures
Similar articles
-
GUN Mutants: New Weapons To Unravel Ascospore Germination Regulation in the Model Fungus Podospora anserina.Microbiol Spectr. 2023 Feb 14;11(2):e0146122. doi: 10.1128/spectrum.01461-22. Online ahead of print. Microbiol Spectr. 2023. PMID: 36786590 Free PMC article.
-
Convergent evolution of morphogenetic processes in fungi: Role of tetraspanins and NADPH oxidases 2 in plant pathogens and saprobes.Commun Integr Biol. 2008;1(2):180-1. doi: 10.4161/cib.1.2.7198. Commun Integr Biol. 2008. PMID: 19704887 Free PMC article.
-
Fungi have three tetraspanin families with distinct functions.BMC Genomics. 2008 Feb 3;9:63. doi: 10.1186/1471-2164-9-63. BMC Genomics. 2008. PMID: 18241352 Free PMC article.
-
Fungal Pls1 tetraspanins as key factors of penetration into host plants: a role in re-establishing polarized growth in the appressorium?FEMS Microbiol Lett. 2006 Mar;256(2):179-84. doi: 10.1111/j.1574-6968.2006.00128.x. FEMS Microbiol Lett. 2006. PMID: 16499604 Review.
-
The genome sequence of Podospora anserina, a classic model fungus.Genome Biol. 2008;9(5):223. doi: 10.1186/gb-2008-9-5-223. Epub 2008 May 15. Genome Biol. 2008. PMID: 18492226 Free PMC article. Review.
Cited by
-
A non-Mendelian MAPK-generated hereditary unit controlled by a second MAPK pathway in Podospora anserina.Genetics. 2012 Jun;191(2):419-33. doi: 10.1534/genetics.112.139469. Epub 2012 Mar 16. Genetics. 2012. PMID: 22426880 Free PMC article.
-
Co-occurrence of tetraspanin and ROS generators: Conservation in protein cross-linking and other developmental processes.Worm. 2013 Apr 1;2(2):e23415. doi: 10.4161/worm.23415. Worm. 2013. PMID: 24058871 Free PMC article.
-
Tpc1 is an important Zn(II)2Cys6 transcriptional regulator required for polarized growth and virulence in the rice blast fungus.PLoS Pathog. 2017 Jul 24;13(7):e1006516. doi: 10.1371/journal.ppat.1006516. eCollection 2017 Jul. PLoS Pathog. 2017. PMID: 28742127 Free PMC article.
-
Appressoria-Small but Incredibly Powerful Structures in Plant-Pathogen Interactions.Int J Mol Sci. 2023 Jan 21;24(3):2141. doi: 10.3390/ijms24032141. Int J Mol Sci. 2023. PMID: 36768468 Free PMC article. Review.
-
The NADPH oxidase in Volvariella volvacea and its differential expression in response to mycelial ageing and mechanical injury.Braz J Microbiol. 2020 Mar;51(1):87-94. doi: 10.1007/s42770-019-00165-4. Epub 2019 Oct 30. Braz J Microbiol. 2020. PMID: 31667800 Free PMC article. Review.
References
-
- Altschul, S. F., W. Gish, W. Miller, E. W. Myers, and D. J. Lipman. 1990. Basic local alignment search tool. J. Mol. Biol. 215403-410. - PubMed
-
- Ausubel, F. M., R. Brent, R. E. Kingston, D. D. Moore, J. G. Seidman, J. A. Smith, and K. Struhl (ed.). 1987. Current protocols in molecular biology. Wiley Interscience, New York, NY.
-
- Beckett, A., R. Barton, and I. M. Wilson. 1968. Fine structure of the wall and appendage formation in ascospores of Podospora anserina. J. Gen. Microbiol. 5389-94. - PubMed
-
- Bedard, K., and K. H. Krause. 2007. The NOX family of ROS-generating NADPH oxidases: physiology and pathophysiology. Physiol. Rev. 87245-313. - PubMed
-
- Bhambra, G. K., Z. Y. Wang, D. M. Soanes, G. E. Wakley, and N. J. Talbot. 2006. Peroxisomal carnitine acetyl transferase is required for elaboration of penetration hyphae during plant infection by Magnaporthe grisea. Mol. Microbiol. 6146-60. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
