

The evolutionary rate of duplicated genes under concerted evolution
- PMID: 18757936
- PMCID: PMC2535699
- DOI: 10.1534/genetics.108.087676
The evolutionary rate of duplicated genes under concerted evolution
Abstract
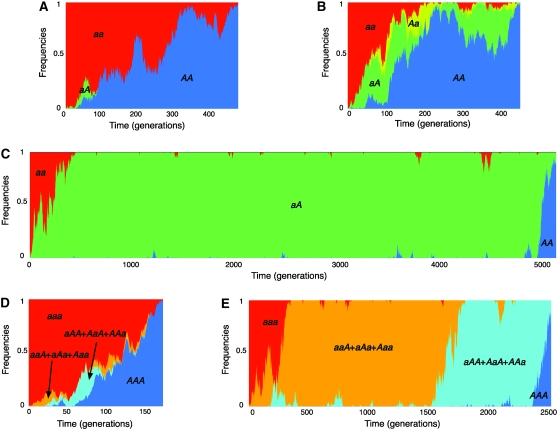
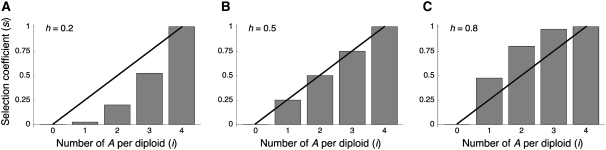
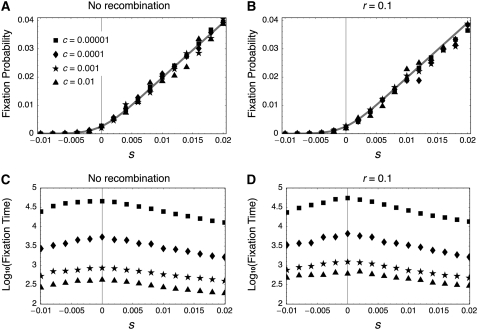
The effect of directional selection on the fixation process of a single mutation that spreads in a multigene family by gene conversion is investigated. A simple two-locus model with two alleles, A and a, is first considered in a random-mating diploid population with size N. There are four haplotypes, AA, Aa, aA, and aa, and selection works on the number of alleles A in a diplod (i = 0, 1, 2, 3, 4). Because gene conversion is allowed between the two loci, when the mutation rate is very low, either AA or aa will fix in the population eventually. We consider a situation where a single mutant, A, arises in one locus when a is fixed in both loci. Then, we derive the fixation probability analytically, and the fixation time is investigated by simulations. It is found that gene conversion has an effect to increase the "effective" population size, so that weak selection works more efficiently in a multigene family. With these results, we discuss the effect of gene conversion on the rate of molecular evolution in a multigene family undergoing concerted evolution. We also argue about the applicability of the theoretical results to models of multigene families with more than two loci.
Figures
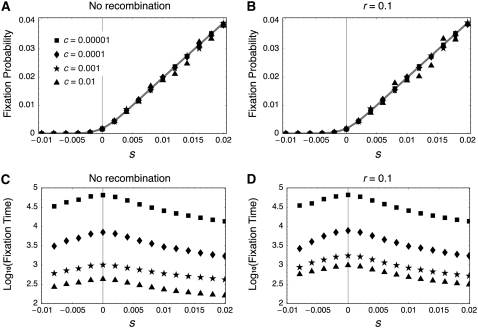
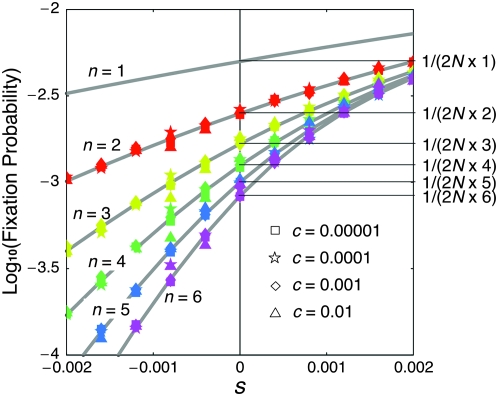
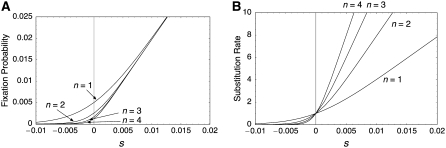
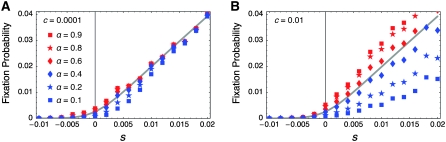
References
-
- Arnheim, N., 1983. Concerted evolution of multigene families, pp. 38–61 in Evolution of Genes and Proteins, edited by M. Nei and R. K. Koehn. Sinauer Associates, Sunderland, MA.
-
- Crow, J. F., and M. Kimura, 1970. An Introduction to Population Genetics Theory. Harper & Row, New York.
-
- Dover, G., 1982. Molecular drive: a cohesive mode of species evolution. Nature 299 111–117. - PubMed
-
- Galtier, N., 2003. Gene conversion drives GC content evolution in mammalian histones. Trends Genet. 19 65–68. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
