

Drosophila nemo promotes eye specification directed by the retinal determination gene network
- PMID: 18757943
- PMCID: PMC2535682
- DOI: 10.1534/genetics.108.092155
Drosophila nemo promotes eye specification directed by the retinal determination gene network
Abstract
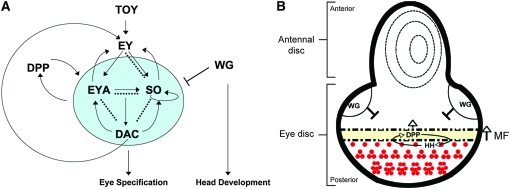
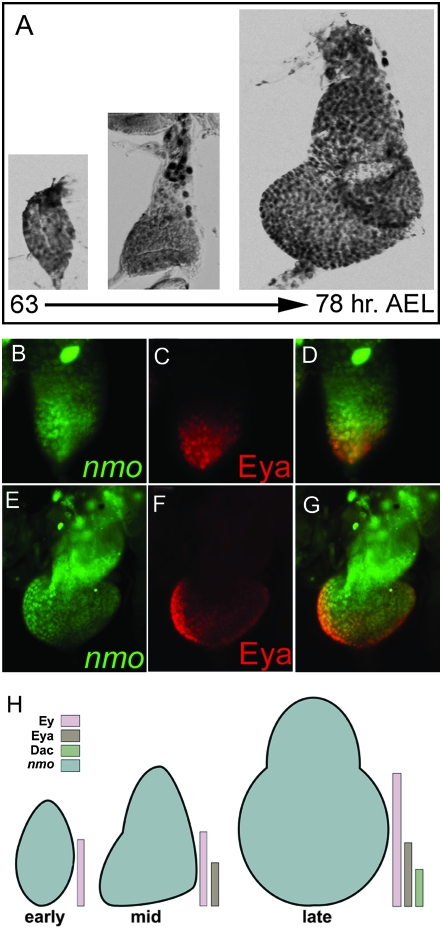
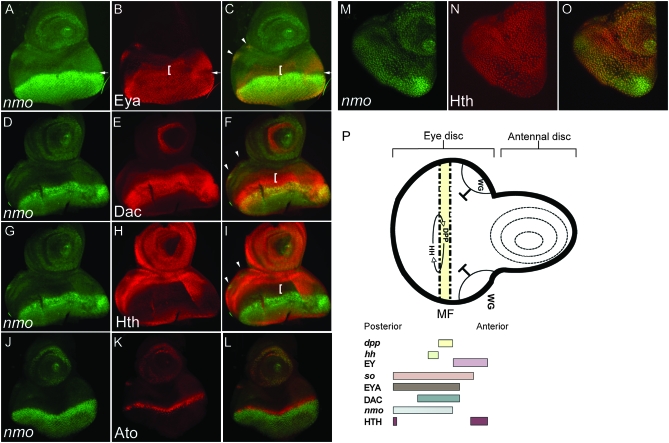
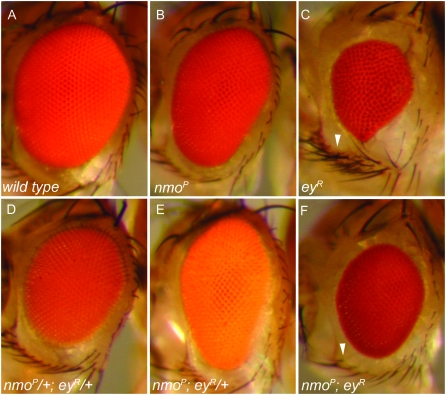
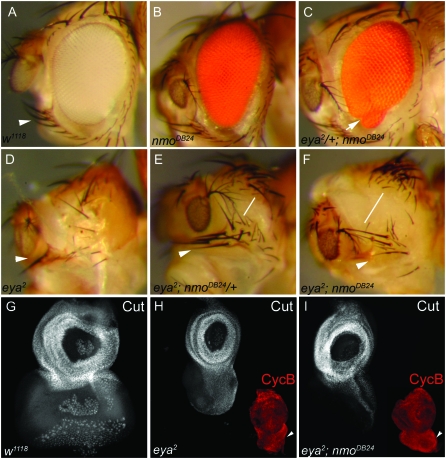
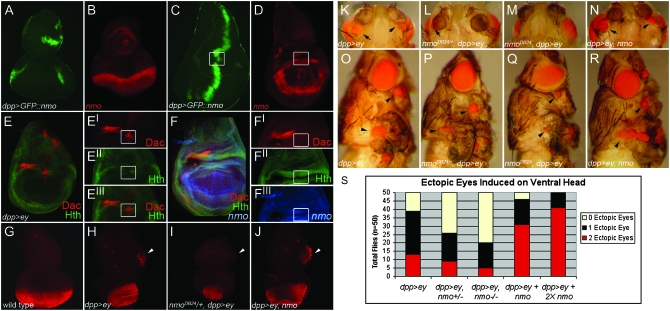
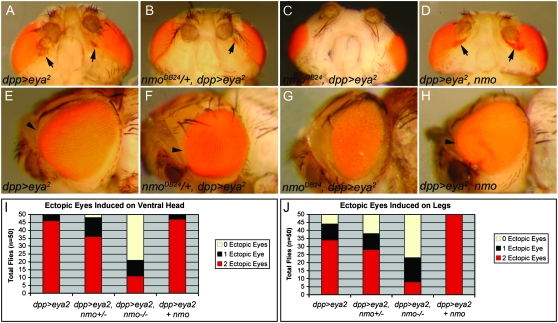
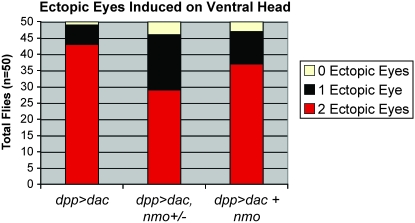
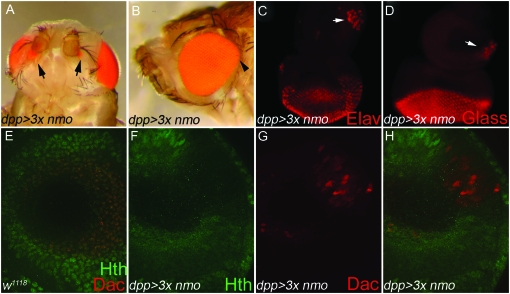
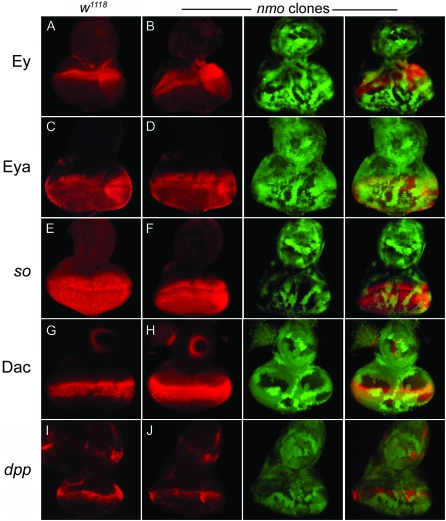
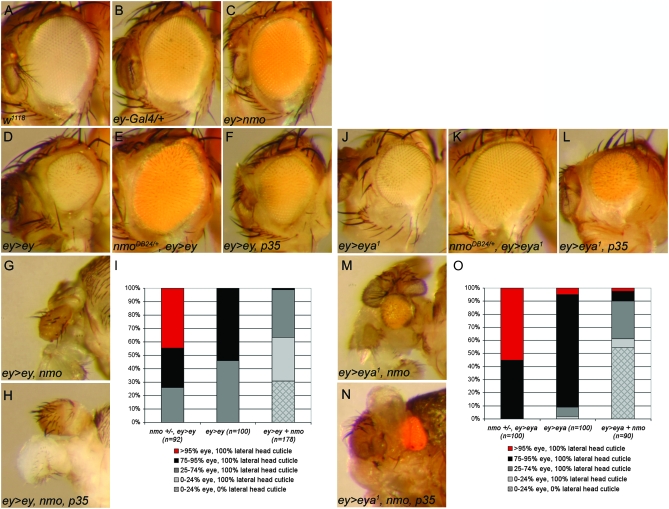
Drosophila nemo (nmo) is the founding member of the Nemo-like kinase (Nlk) family of serine-threonine kinases. Previous work has characterized nmo's role in planar cell polarity during ommatidial patterning. Here we examine an earlier role for nmo in eye formation through interactions with the retinal determination gene network (RDGN). nmo is dynamically expressed in second and third instar eye imaginal discs, suggesting additional roles in patterning of the eyes, ocelli, and antennae. We utilized genetic approaches to investigate Nmo's role in determining eye fate. nmo genetically interacts with the retinal determination factors Eyeless (Ey), Eyes Absent (Eya), and Dachshund (Dac). Loss of nmo rescues ey and eya mutant phenotypes, and heterozygosity for eya modifies the nmo eye phenotype. Reducing nmo also rescues small-eye defects induced by misexpression of ey and eya in early eye development. nmo can potentiate RDGN-mediated eye formation in ectopic eye induction assays. Moreover, elevated Nmo alone can respecify presumptive head cells to an eye fate by inducing ectopic expression of dac and eya. Together, our genetic analyses reveal that nmo promotes normal and ectopic eye development directed by the RDGN.
Figures
Similar articles
-
Nemo phosphorylates Eyes absent and enhances output from the Eya-Sine oculis transcriptional complex during Drosophila retinal determination.Dev Biol. 2012 May 1;365(1):267-76. doi: 10.1016/j.ydbio.2012.02.030. Epub 2012 Feb 25. Dev Biol. 2012. PMID: 22394486 Free PMC article.
-
Nemo regulates cell dynamics and represses the expression of miple, a midkine/pleiotrophin cytokine, during ommatidial rotation.Dev Biol. 2013 May 1;377(1):113-25. doi: 10.1016/j.ydbio.2013.02.006. Epub 2013 Feb 18. Dev Biol. 2013. PMID: 23428616 Free PMC article.
-
Hipk promotes photoreceptor differentiation through the repression of Twin of eyeless and Eyeless expression.Dev Biol. 2014 Jun 1;390(1):14-25. doi: 10.1016/j.ydbio.2014.02.024. Epub 2014 Mar 12. Dev Biol. 2014. PMID: 24631217
-
Retinal determination the beginning of eye development.Curr Top Dev Biol. 2010;93:1-28. doi: 10.1016/B978-0-12-385044-7.00001-1. Curr Top Dev Biol. 2010. PMID: 20959161 Free PMC article. Review.
-
[Constructive synergism of regulatory genes expressed in the course of the eye and muscle development and regeneration].Izv Akad Nauk Ser Biol. 2001 May-Jun;(3):261-75. Izv Akad Nauk Ser Biol. 2001. PMID: 11433936 Review. Russian.
Cited by
-
The fly eye: Through the looking glass.Dev Dyn. 2018 Jan;247(1):111-123. doi: 10.1002/dvdy.24585. Epub 2017 Oct 23. Dev Dyn. 2018. PMID: 28856763 Free PMC article. Review.
-
Nemo phosphorylates Eyes absent and enhances output from the Eya-Sine oculis transcriptional complex during Drosophila retinal determination.Dev Biol. 2012 May 1;365(1):267-76. doi: 10.1016/j.ydbio.2012.02.030. Epub 2012 Feb 25. Dev Biol. 2012. PMID: 22394486 Free PMC article.
-
My what big eyes you have: how the Drosophila retina grows.Dev Neurobiol. 2011 Dec;71(12):1133-52. doi: 10.1002/dneu.20921. Dev Neurobiol. 2011. PMID: 21604387 Free PMC article. Review.
-
Identification of novel components of the retinal determination gene network in Drosophila cell lines.Open Biol. 2025 Jul;15(7):250012. doi: 10.1098/rsob.250012. Epub 2025 Jul 16. Open Biol. 2025. PMID: 40664237 Free PMC article.
-
Retinal Expression of the Drosophila eyes absent Gene Is Controlled by Several Cooperatively Acting Cis-regulatory Elements.PLoS Genet. 2016 Dec 8;12(12):e1006462. doi: 10.1371/journal.pgen.1006462. eCollection 2016 Dec. PLoS Genet. 2016. PMID: 27930646 Free PMC article.
References
-
- Azpiazu, N., and G. Morata, 2000. Function and regulation of homothorax in the wing imaginal disc of Drosophila. Development 127 2685–2693. - PubMed
-
- Azpiazu, N., and G. Morata, 2002. Distinct functions of homothorax in leg development in Drosophila. Mech. Dev. 119 55–67. - PubMed
-
- Baonza, A., and M. Freeman, 2002. Control of Drosophila eye specification by Wingless signalling. Development 129 5313–5322. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
