

Heterologous ectoine production in Escherichia coli: by-passing the metabolic bottle-neck
- PMID: 18759971
- PMCID: PMC2562377
- DOI: 10.1186/1746-1448-4-12
Heterologous ectoine production in Escherichia coli: by-passing the metabolic bottle-neck
Abstract
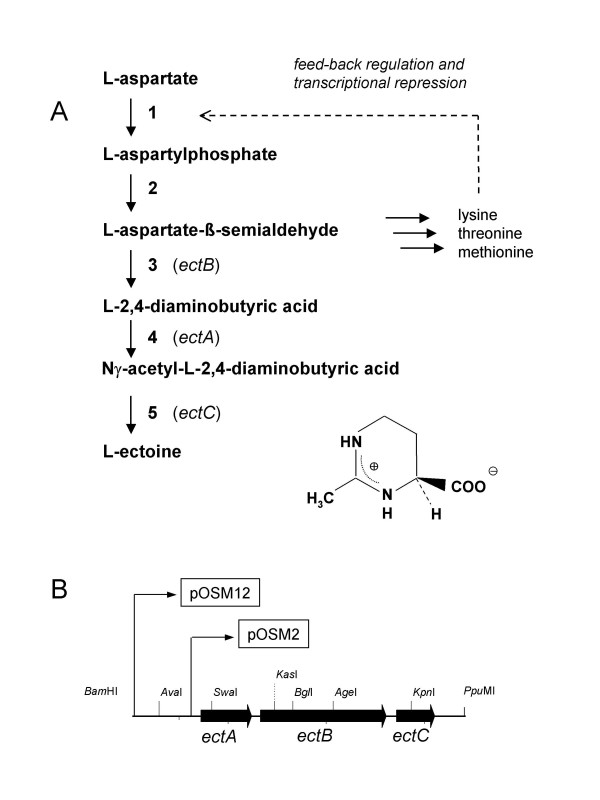
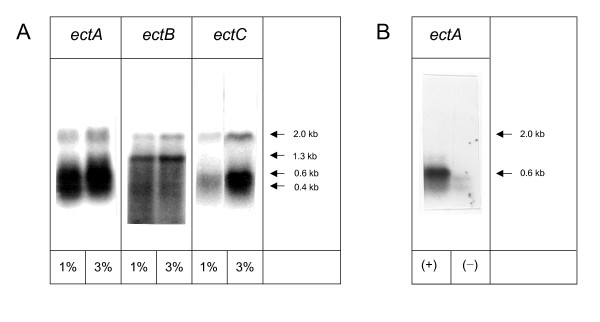
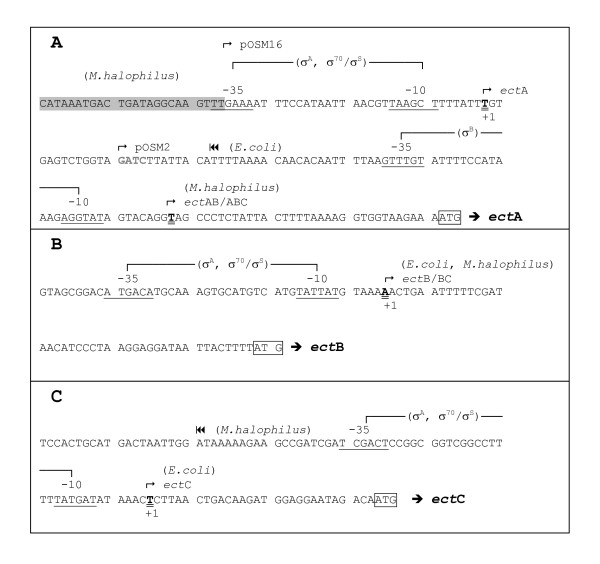
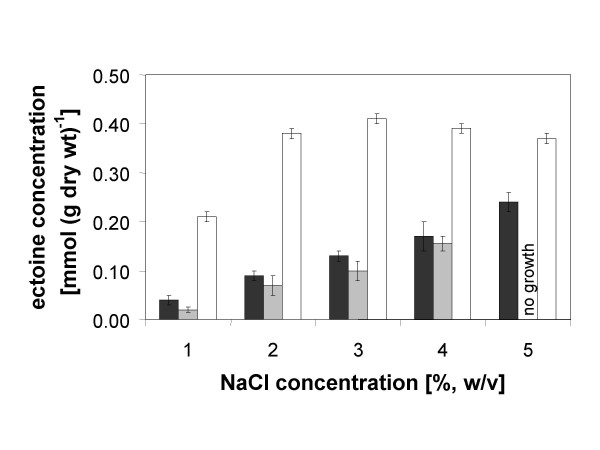
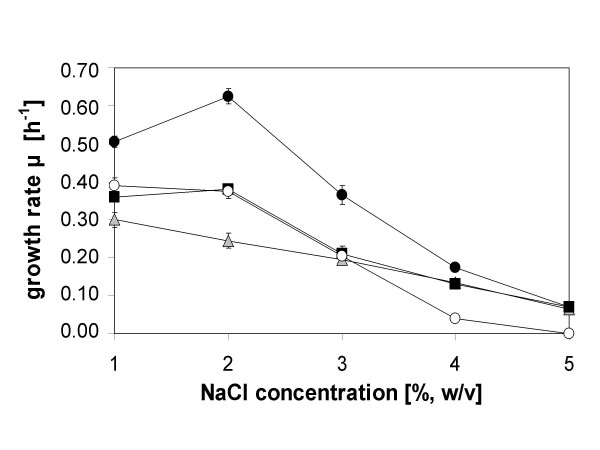
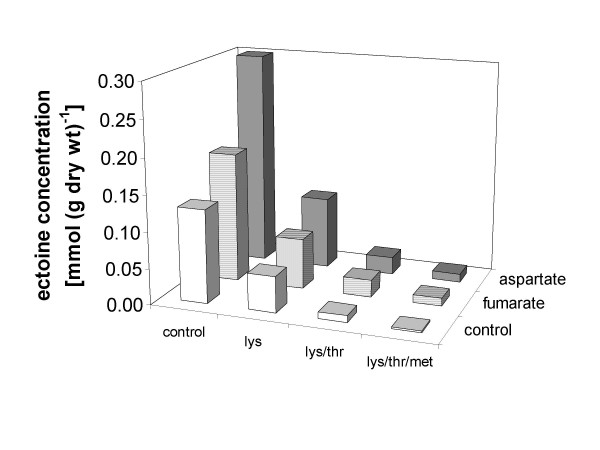
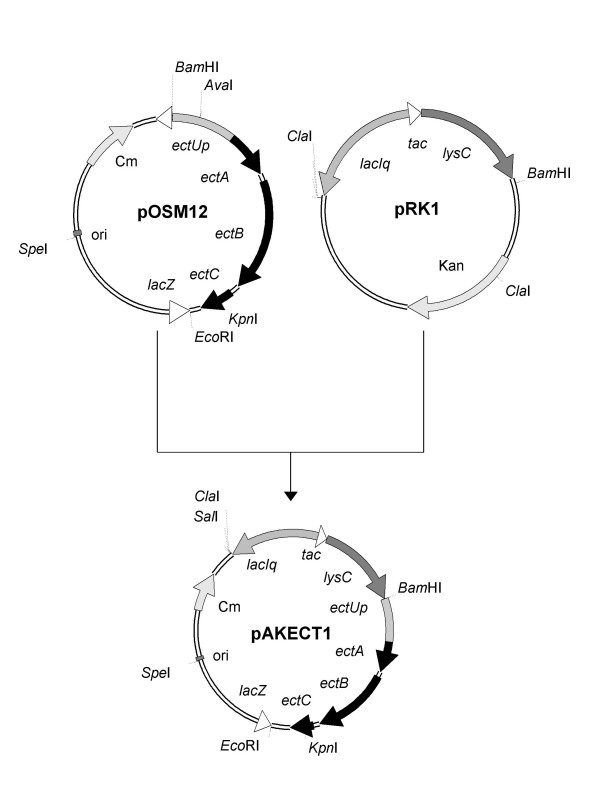
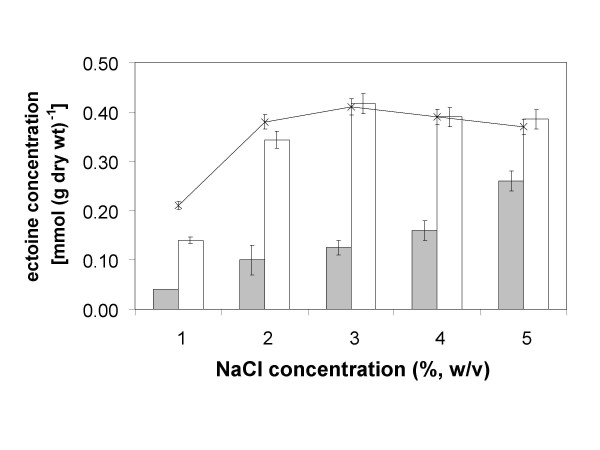
Transcription of the ectoine biosynthesis genes ectA, ectB and ectC from Marinococcus halophilus in recombinant Escherichia coli DH5alpha is probably initiated from three individual sigma70/sigmaA-dependent promoter sequences, upstream of each gene. Consequently, mRNA-fragments containing the single genes and combinations of the genes ectA and ectB or ectB and ectC, respectively, could be detected by Northern blot analysis. Under the control of its own regulatory promoter region (ectUp) a seemingly osmoregulated ectoine production was observed. In addition, aspartate kinases were identified as the main limiting factor for ectoine production in recombinant E. coli DH5alpha. Co-expression of the ectoine biosynthesis genes and of the gene of the feedback-resistant aspartate kinase from Corynebacterium glutamicum MH20-22B (lysC) led to markedly increased production of ectoine in E. coli DH5alpha, resulting in cytoplasmic ectoine concentrations comparable to those reached via ectoine accumulation from the medium.
Figures
References
-
- da Costa MS, Santos H, Galinski EA. An overview of the role and diversity of compatible solutes in bacteria and archaea. Adv Biochem Eng Biotechnol. 1998;61:117–153. - PubMed
-
- Bremer E, Krämer R. In: Bacterial Stress Responses. Storz G, Hengge-Aronis R, editor. Washington D.C.: ASM Press; 2000. Coping with osmotic challenges: osmoregulation through accumulation and release of compatible solutes; pp. 79–97.
-
- Galinski EA, Pfeiffer HP, Trüper HG. 1,4,5,6-Tetrahydro-2-methyl-4-pyrimidine carboxylic acid: A novel cyclic amino acid from halophilic phototrophic bacteria of the genus Ectothiorhodospira. Eur J Biochem. 1985;149:135–139. - PubMed
-
- Severin J, Wohlfahrt A, Galinski EA. The predominant role of recently discovered tetrahydropyrimidines for the osmoadaptation of halophilic eubacteria. J Gen Microbiol. 1992;138:1629–1638.
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
