

Folding mechanism of reduced Cytochrome c: equilibrium and kinetic properties in the presence of carbon monoxide
- PMID: 18761351
- PMCID: PMC2653224
- DOI: 10.1016/j.jmb.2008.08.025
Folding mechanism of reduced Cytochrome c: equilibrium and kinetic properties in the presence of carbon monoxide
Abstract
Despite close structural similarity, the ferric and ferrous forms of cytochrome c differ greatly in terms of their ligand binding properties, stability, folding, and dynamics. The reduced heme iron binds diatomic ligands such as CO only under destabilizing conditions that promote weakening or disruption of native methionine-iron linkage. This makes CO a useful conformational probe for detecting partially structured states that cannot be observed in the absence of endogenous ligands. Heme absorbance, circular dichroism, and NMR were used to characterize the denaturant-induced unfolding equilibrium of ferrocytochrome c in the presence and in the absence of CO. In addition to the native state (N), which does not bind CO, and the unfolded CO complex (U-CO), a structurally distinct CO-bound form (M-CO) accumulates to high levels (approximately 75% of the population) at intermediate guanidine HCl concentrations. Comparison of the unfolding transitions for different conformational probes reveals that M-CO is a compact state containing a native-like helical core and regions of local disorder in the segment containing the native Met80 ligand and adjacent loops. Kinetic measurements of CO binding and dissociation under native, partially denaturing, and fully unfolded conditions indicate that a state M that is structurally analogous to M-CO is populated even in the absence of CO. The binding energy of the CO ligand lowers the free energy of this high-energy state to such an extent that it accumulates even under mildly denaturing equilibrium conditions. The thermodynamic and kinetic parameters obtained in this study provide a fully self-consistent description of the linked unfolding/CO binding equilibria of reduced cytochrome c.
Figures




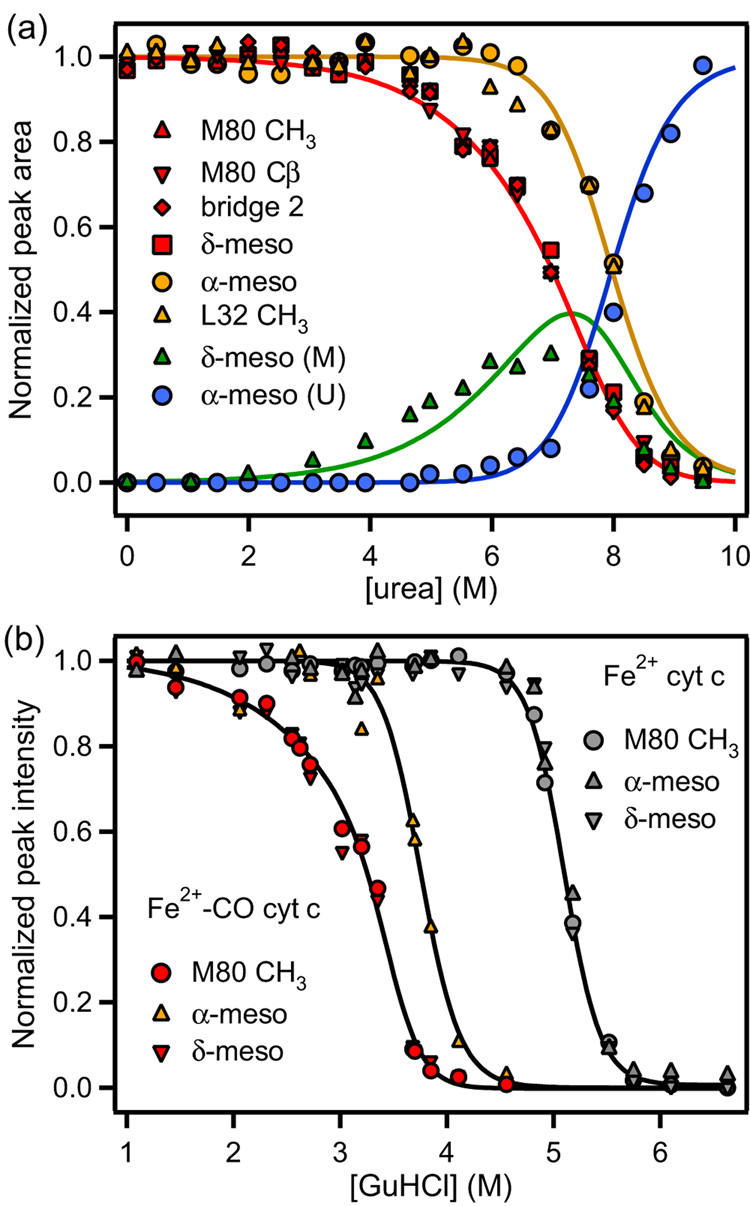
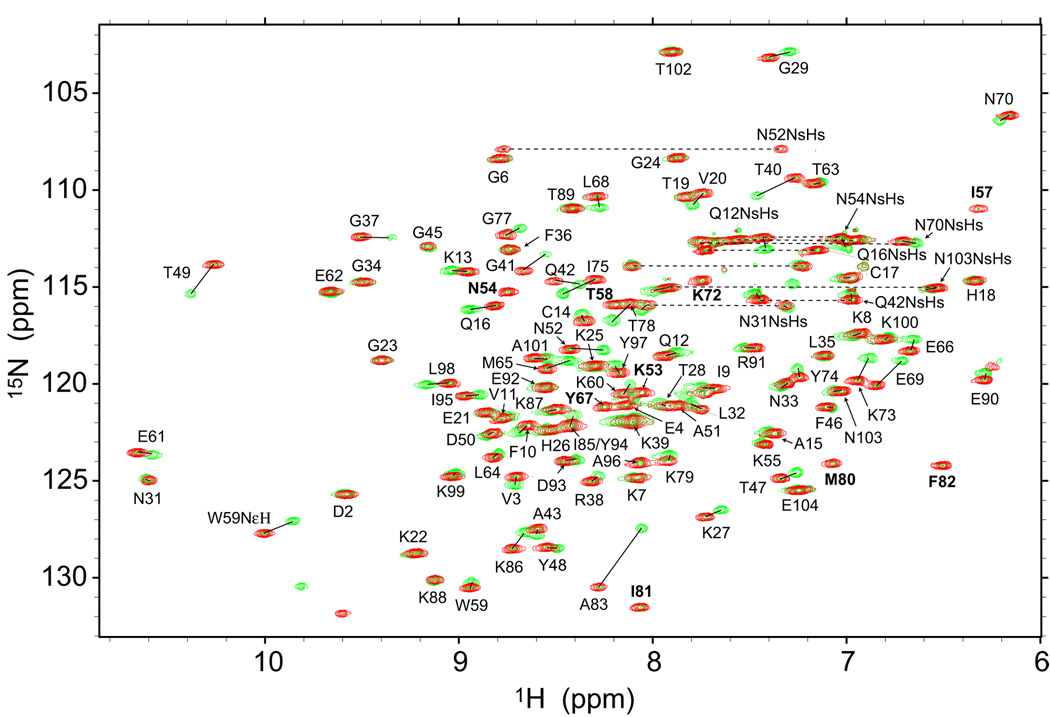



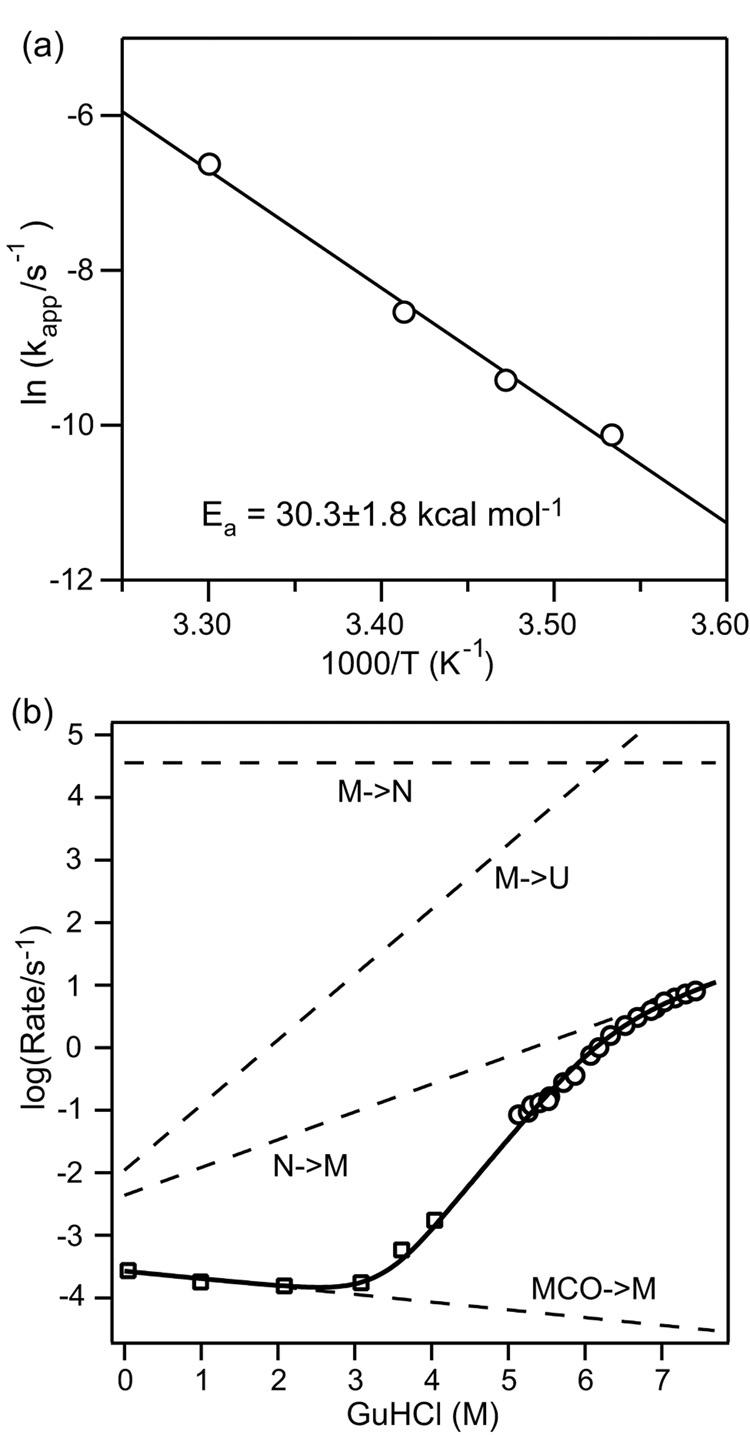



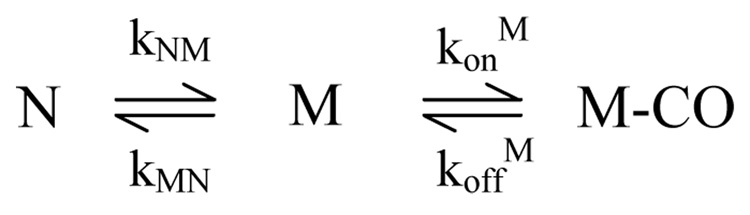
References
-
- Feng YQ, Wand AJ, Sligar SG. 1H and 15N NMR resonance assignments and preliminary structural characterization of Escherichia coli apocytochrome b562. Biochemistry. 1991;30:7711–7717. - PubMed
-
- Stellwagen E, Rysavy R, Babul G. The conformation of horse heart apocytochrome c. J. Biol. Chem. 1972;247:8074–8077. - PubMed
-
- Fisher WR, Taniuchi H, Anfinsen CB. On the role of heme in the formation of the structure of cytochrome c. J. Biol. Chem. 1973;248:3188–3195. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
