

Review
doi: 10.1016/j.dci.2008.07.011.
Epub 2008 Aug 30.
The ontogeny of the porcine immune system
Affiliations
- PMID: 18762210
- PMCID: PMC7103207
- DOI: 10.1016/j.dci.2008.07.011
Item in Clipboard
Review
The ontogeny of the porcine immune system
Dev Comp Immunol.
2009 Mar.
Abstract
Cellular and humoral aspects of the immune response develop sequentially in the fetus. During the ontogeny, the pluripotent stem cells emerge and differentiate into all hematopoietic lineages. Basic questions including the identification of the first lympho-hematopoietic sites, the origin of T and B lymphocytes, the development of different subpopulations of alphabeta T, gammadelta T and B lymphocytes as well as development of innate immunity and the acquisition of full immunological capacities are discussed here for swine and compared with other species. The description of related topics such as fertilization, morphogenesis, maternal-fetal-neonatal physiology and early neonatal development are also discussed.
Figures
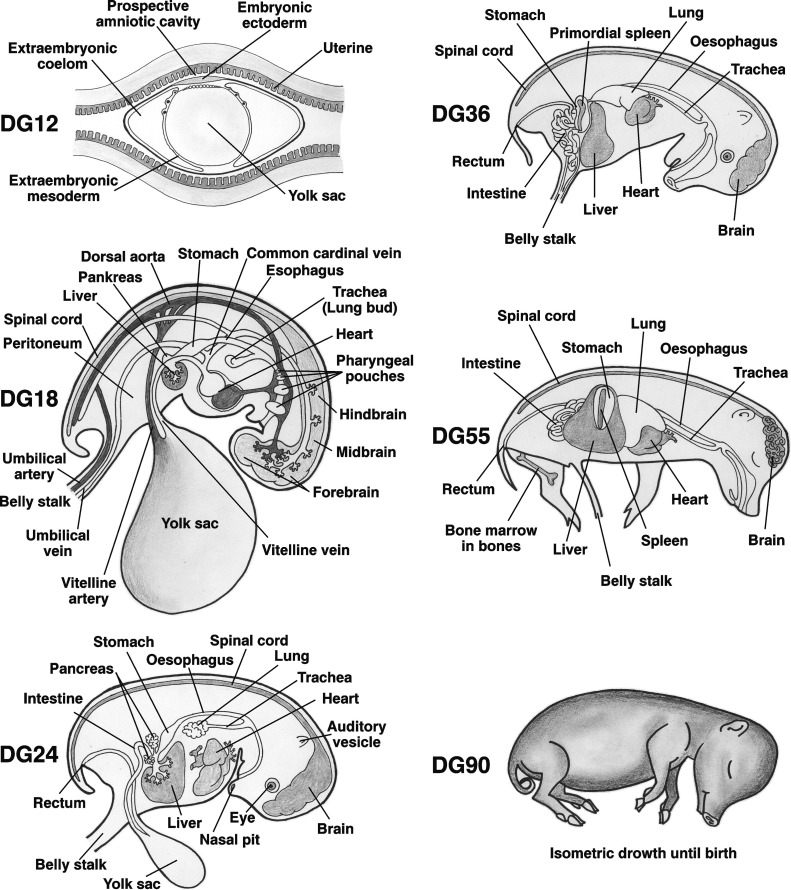
Schematic illustration of fetal development of porcine organs.
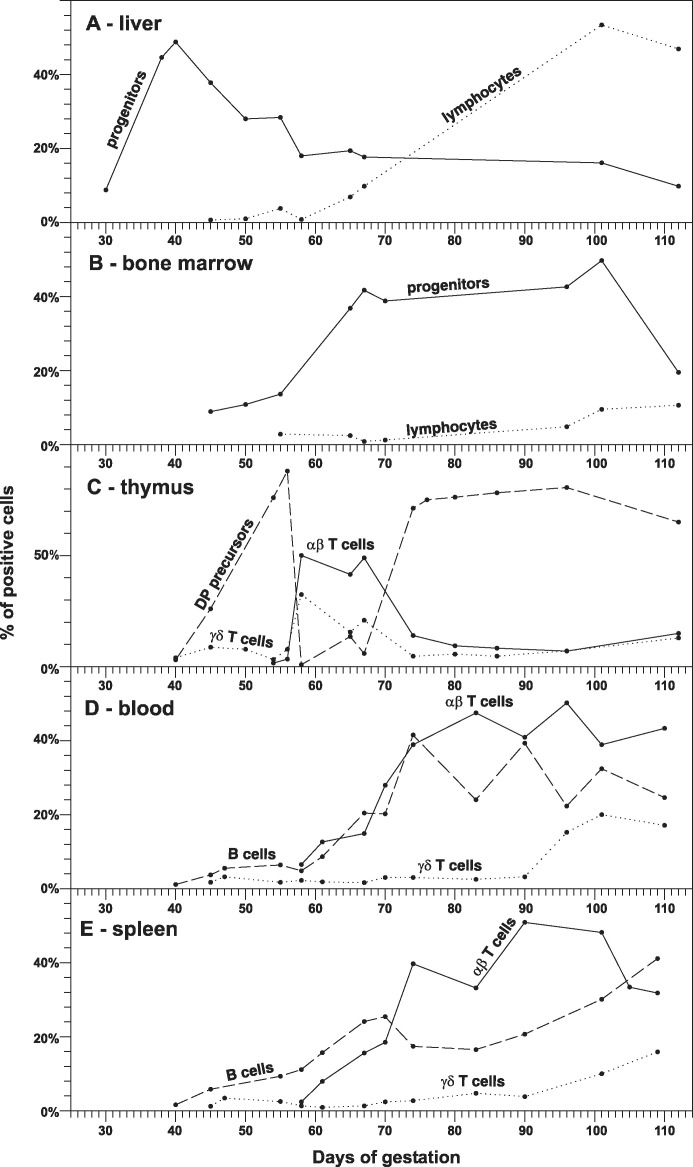
The ontogeny of primary lymphopoietic activity in fetal liver (A), bone marrow (B) and thymus (C). The proportions of putative lymphoid progenitors (CD45loCD172a− cells, solid line) and mature lymphocytes (CD45hiCD172a− cells, dotted line) in fetal liver (A) and bone marrow (B) demonstrate the shift of lymphopoietic activity between these hematopoietic centers. The thymus (C) is populated by pro-T cell progenitors from the same hematopoietic centers that can be easily observed by two waves of developing immature DP precursors (large CD4+CD8+ cells that are CD3−TCRαβ−TCRγδ−, dashed line). Relationship to the development of mature αβ (small CD3hiTCRαβ+ cells, solid line) and γδ (small CD3hiTCRγδ+ cells, dotted line) T lymphocytes in the thymus is also shown. The ontogeny of occurrence for peripheral mature αβ T cells (solid line), γδ T cells (dotted line) and B cells (dashed line) is shown for fetal blood (D) and spleen (E). Data are based on results previously published by Sinkora et al. , , , , .
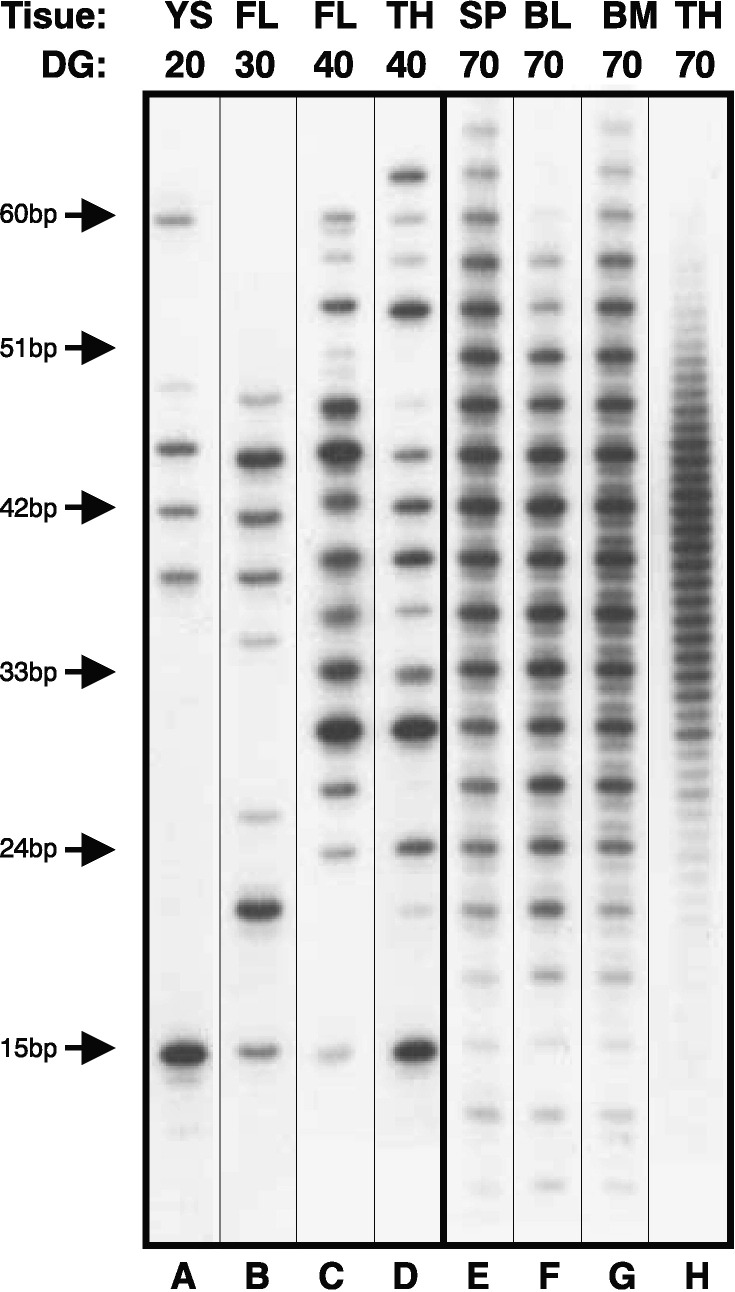
Length analysis of the HCDR3 repertoire recovered from DNA amplifications of VDJH in representative samples coming from yolk sac (YS), fetal liver (FL), thymus (TH), spleen (SP), blood (BL) and bone marrow (BM) during different days of gestation (DG). The left panels (A–D) represent samples recovered from fetuses before the bone marrow starts its hemapoietic and lymphopoietic activity (before at about DG45) while right panels (E–H) represent samples isolated from fetuses after the bone marrow become active. HCDR3 length markers are indicated on the left. Note that HCDR3s isolated from B cells coming from the first wave of progenitors (panels A–D) present a pattern in which there are no out-of-frame rearrangements (no minor bands in lengths between in-frame rearrangements). On the other hand, B cells arising from the second wave of progenitors that originate in the bone marrow (panels E–G) contain out-of-frame rearrangements that represent non-productive rearrangements. The VDJH rearrangements isolated from thymus after DG55 (panel H) displays the total absence of selection for in-frame rearrangements.
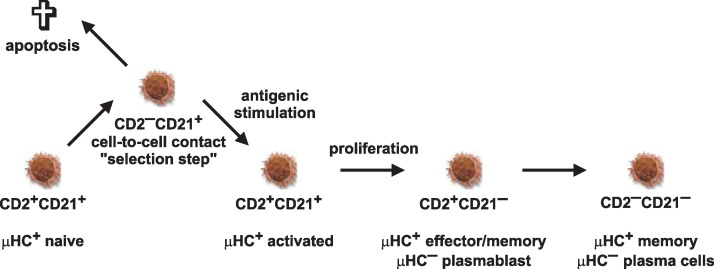
Proposed model of peripheral B cell maturation and diversification. CD2+CD21+ B cells represent naive mature B cells that down-regulate CD2 following cell-to-cell contact and activation. A portion of the activated CD2−CD21+ B cells probably died while survivors re-establish CD2 expression to become again CD2+CD21+. Further maturation of activated B cells includes down-regulation of CD21. Resultant CD2+CD21− B cells can subsequently generate μHC− large proliferating plasmablasts or small non-dividing plasma cells that eventually become CD2−CD21−. This schematic representation is deduced from recent findings , , . The phenotype and functional status of individual subsets of B cells is given.
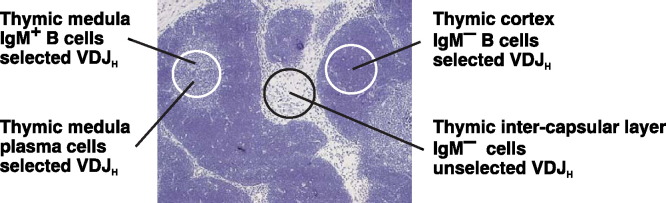
The localization of thymic cell populations that have rearranged their immunoglobulin VDJH genes. One population can be found exclusively in thymic medulla and consists mainly of immature and mature sIgM+ B cells and plasma cells, all of which have productively rearranged their VDJH genes. The second population found mainly in thymic cortex are sIgM− cells that possess characteristics of a selected population, similar to B cells arising from the bone marrow. The third population is found in the thymic inter-capsular layer and shows no evidence of selection for in-frame VDJH rearrangement. These cells do not express a BCR and do not bear other known swine B cell surface markers.
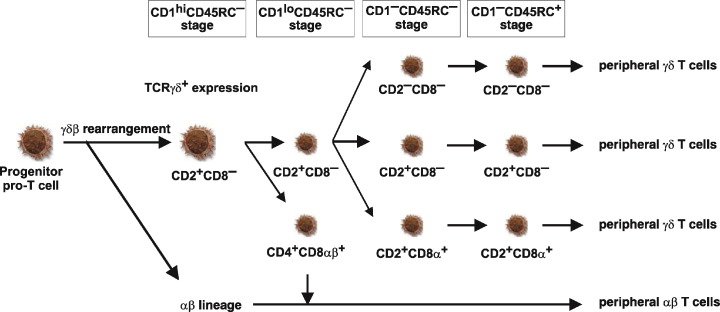
Proposed model of γδ T cell development and diversification in the thymus. Large cell icons demonstrate differentiation stages of large, mitotically active thymocytes while small cell icons represent small, non-dividing populations. The phenotype of individual subsets is given. Developmental stages determined using CD1 and CD45RC expression are summarized at the top. This schematic representation is deduced from recent findings , .
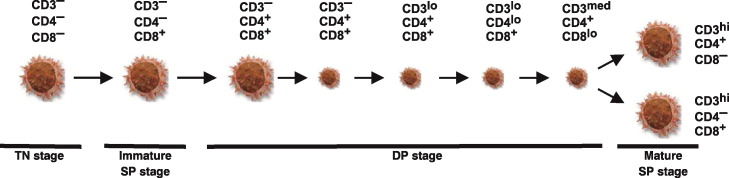
Proposed model of αβ T cell development. Large cell icons demonstrate differentiation stages of large, mitotically active thymocytes while small cell icons represent small, non-dividing populations. The phenotype of individual subsets is given. This schematic representation is deduced from recent findings .
References
-
- Marrable A.W. Pitman Medical Publishing; Great Britain: 1971. The embryonic pig: a chronological account.
-
- Sterzl J., Silverstein A.M. Developmental aspects of immunity. Adv Immunol. 1967;6(1):337–371. - PubMed
-
- Butler J.E., Brown W.R. The immunoglobulins and immunoglobulin genes of swine. Vet Immunol Immunopathol. 1994;43(1-3):5–12. - PubMed
-
- Tlaskalova-Hogenova H., Mandel L., Trebichavsky I., Kovaru F., BArot R., Sterzl J. Development of immune responses in early pig ontogeny. Vet Immunol Immunopathol. 1994;43(1-3):135–142. - PubMed
-
- Butler J.E., Sun J., Weber P., Ford S.P., Rehakova Z., Sinkora J. Antibody repertoire development in fetal and neonatal piglets. IV. Switch recombination, primarily in fetal thymus occurs independent of environmental antigen and is only weakly associated with repertoire diversification. J Immunol. 2001;167(6):3239–3249. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
