

IGFBP-5 regulates muscle cell differentiation by binding to IGF-II and switching on the IGF-II auto-regulation loop
- PMID: 18762576
- PMCID: PMC2528583
- DOI: 10.1083/jcb.200712110
IGFBP-5 regulates muscle cell differentiation by binding to IGF-II and switching on the IGF-II auto-regulation loop
Abstract
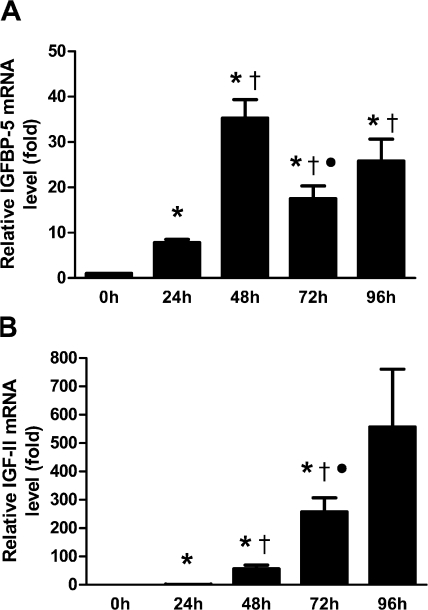
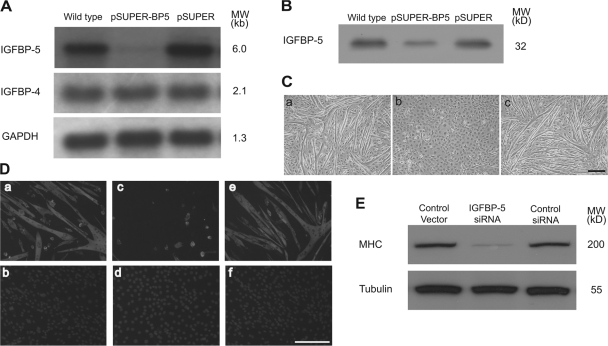
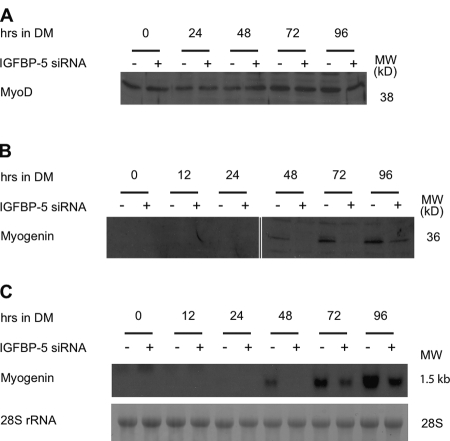
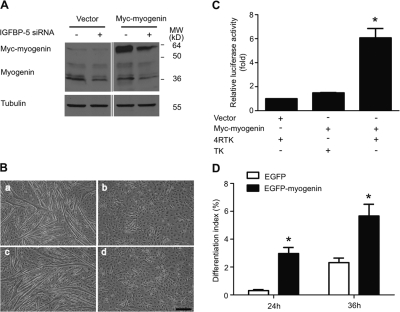
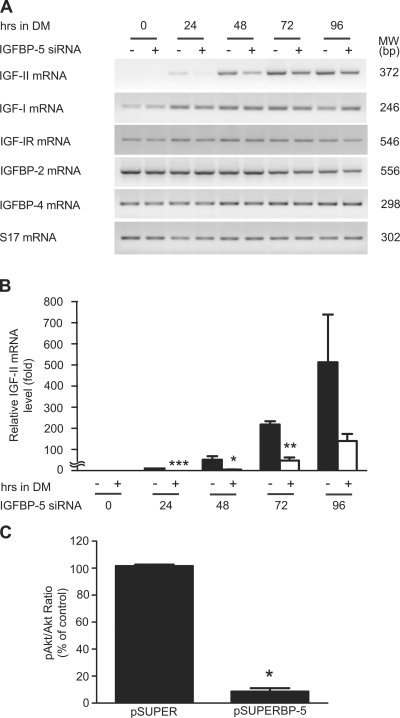
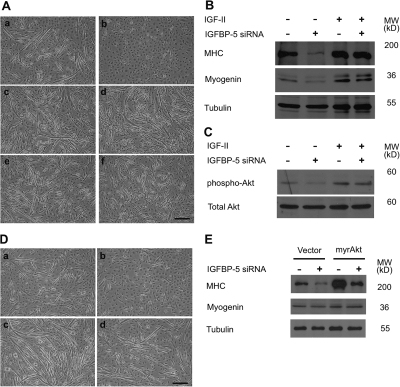
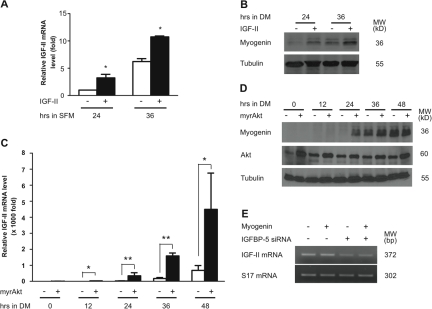
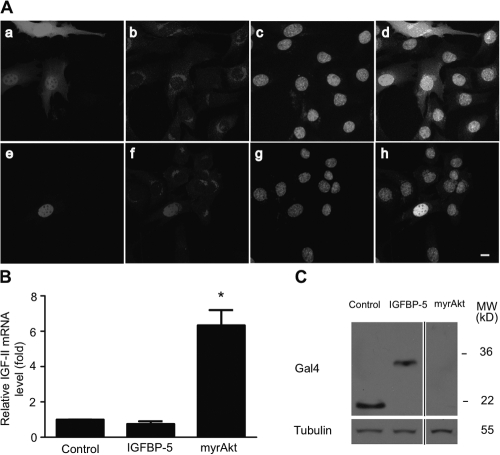
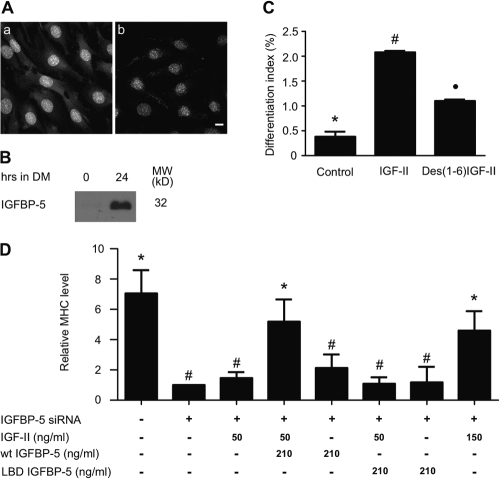
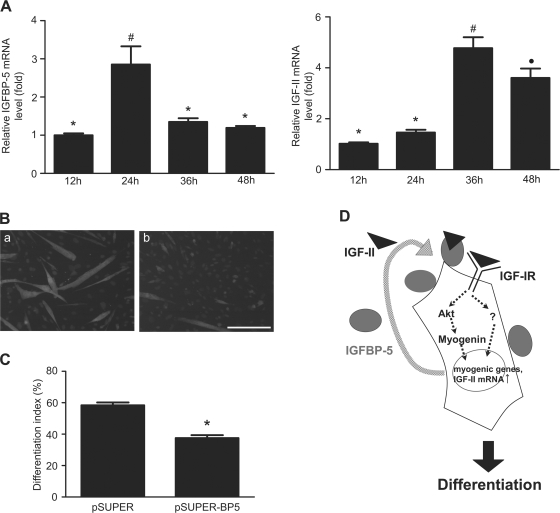
IGF-II stimulates both mitogenesis and myogenesis through its binding and activation of the IGF-I receptor (IGF-IR). How this growth factor pathway promotes these two opposite cellular responses is not well understood. We investigate whether local IGF binding protein-5 (IGFBP-5) promotes the myogenic action of IGF-II. IGFBP-5 is induced before the elevation of IGF-II expression during myogenesis. Knockdown of IGFBP-5 impairs myogenesis and suppresses IGF-II gene expression. IGF-II up-regulates its own gene expression via the PI3K-Akt signaling pathway. Adding IGF-II or constitutively activating Akt rescues the IGFBP-5 knockdown-caused defects. However, an IGF analogue that binds to the IGF-IR but not IGFBP has only a limited effect. When added with low concentrations of IGF-II, IGFBP-5 restores IGF-II expression and myogenic differentiation, whereas an IGF binding-deficient IGFBP-5 mutant has no effect. These findings suggest that IGFBP-5 promotes muscle cell differentiation by binding to and switching on the IGF-II auto-regulation loop.
Figures
References
-
- Bayol, S., P.T. Loughna, and C. Brownson. 2000. Phenotypic expression of IGF binding protein transcripts in muscle, in vitro and in vivo. Biochem. Biophys. Res. Commun. 273:282–286. - PubMed
-
- Boutinaud, M., J.H. Shand, M.A. Park, K. Phillips, J. Beattie, D.J. Flint, and G.J. Allan. 2004. A quantitative RT-PCR study of the mRNA expression profile of the IGF axis during mammary gland development. J. Mol. Endocrinol. 33:195–207. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
