

Smad7 is inactivated through a direct physical interaction with the LIM protein Hic-5/ARA55
- PMID: 18762808
- PMCID: PMC3285457
- DOI: 10.1038/onc.2008.291
Smad7 is inactivated through a direct physical interaction with the LIM protein Hic-5/ARA55
Erratum in
-
Correction to: Smad7 is inactivated through a direct physical interaction with the LIM protein Hic-5/ARA55.Oncogene. 2022 Dec;41(50):5411-5412. doi: 10.1038/s41388-022-02510-8. Oncogene. 2022. PMID: 36280700 No abstract available.
Abstract
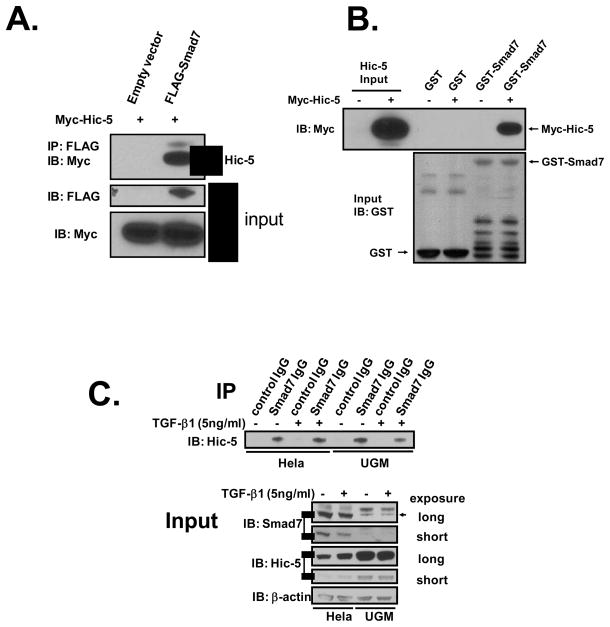
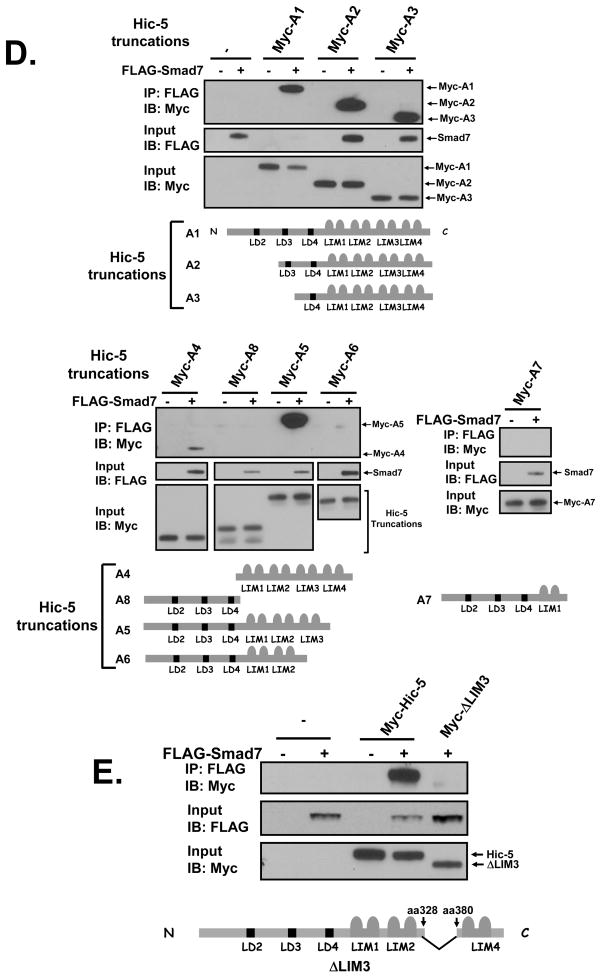
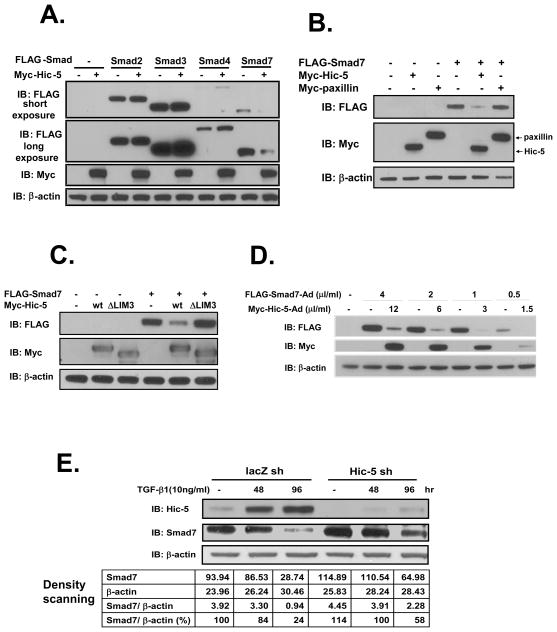
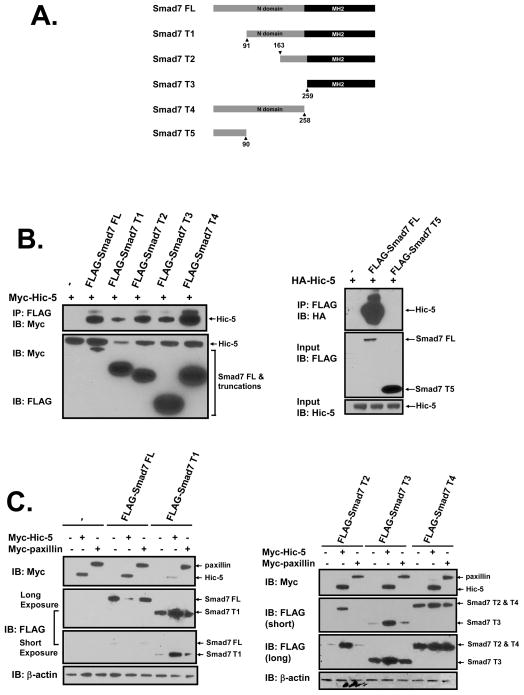
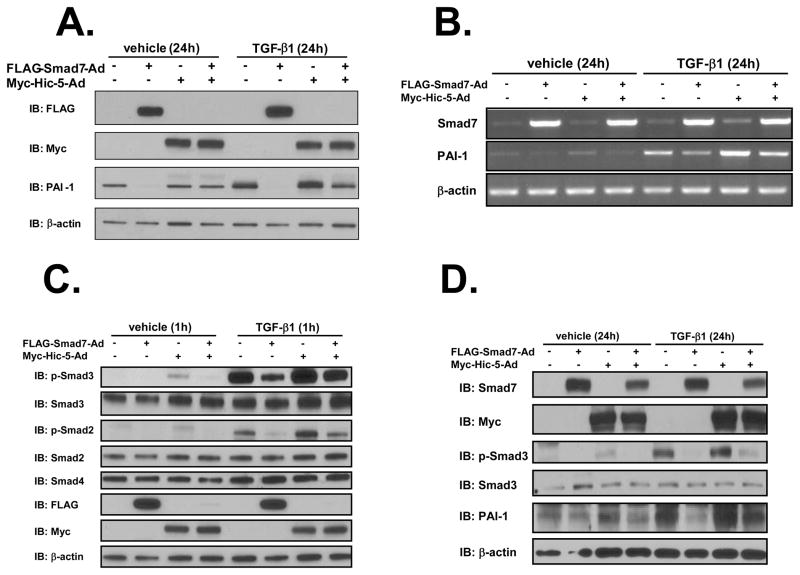
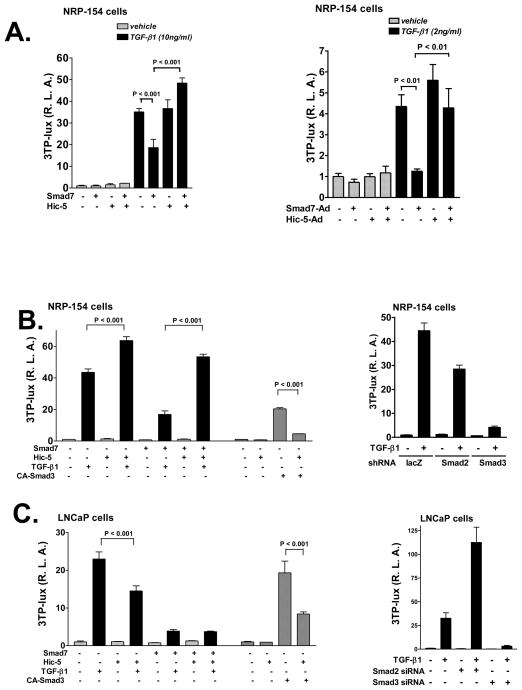
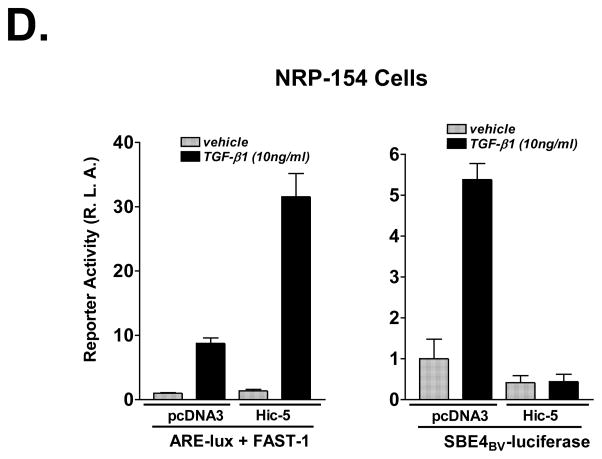
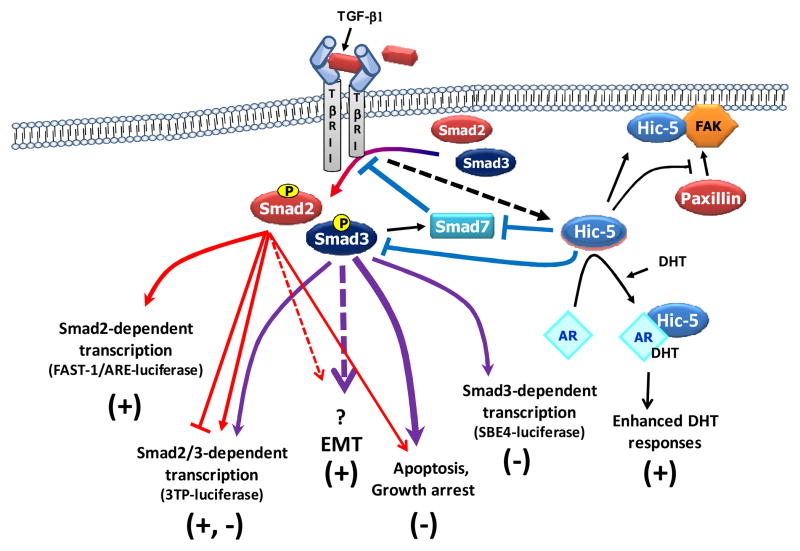
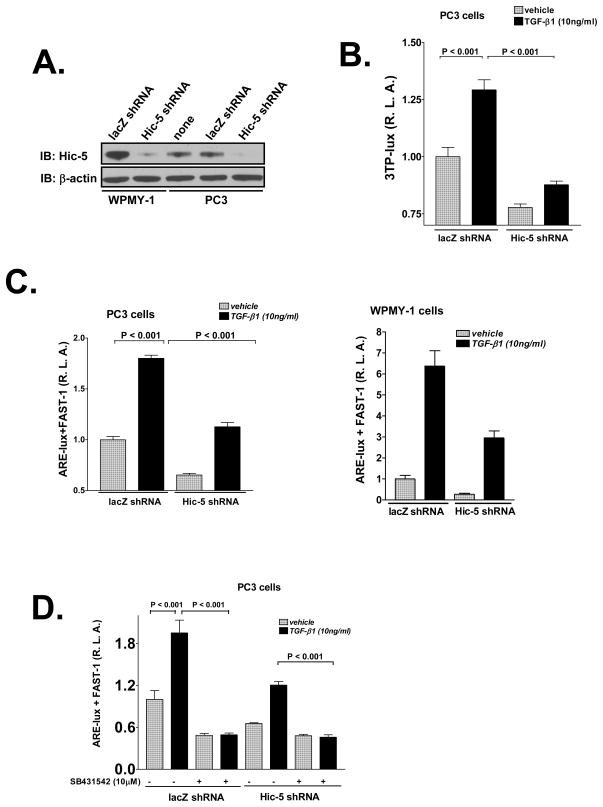
We recently reported that hydrogen peroxide-inducible clone-5 (Hic-5, also named androgen receptor-associated protein 55) can bind to the transforming growth factor-beta (TGF-beta)-signaling regulator Smad3, thereby inhibiting certain Smad3-dependent TGF-beta responses. We now show that Hic-5 can also control TGF-beta responses through an alternative mechanism involving Smad7, a key negative regulator of TGF-beta signaling. Hic-5 binds directly to Smad7. This interaction requires the LIM3 domain of Hic-5, and enhances TGF-beta signaling through causing loss of Smad7 protein but not mRNA. Enforced expression of Hic-5 reverses the ability of Smad7 to suppress TGF-beta-induced phosphorylation of Smads 2 and 3 and activation of the plasminogen activator inhibitor-1 promoter (in NRP-154 and PC3 prostate carcinoma and WPMY-1 prostate myofibroblast cell lines). Lentiviral-mediated small-hairpin RNA silencing of endogenous Hic-5 reduced TGF-beta responses in PC3 and WPMY-1 cells. Further work suggests that the level of Smad7 is modulated by its physical interaction with Hic-5 and targeted to a degradation pathway not likely to be proteasomal. Our findings support that Hic-5 functions as a cell-type-specific activator of TGF-beta signaling through its ability to physically interact with and neutralize Smad7.
Figures
References
-
- Chen X, Weisberg E, Fridmacher V, Watanabe M, Naco G, Whitman M. Smad4 and FAST-1 in the assembly of activin-responsive factor. Nature. 1997;389:85–9. - PubMed
-
- Chipuk JE, Cornelius SC, Pultz NJ, Jorgensen JS, Bonham MJ, Kim SJ, et al. The androgen receptor represses transforming growth factor-beta signaling through interaction with Smad3. J Biol Chem. 2002a;277:1240–8. - PubMed
-
- Chipuk JE, Stewart LV, Ranieri A, Song K, Danielpour D. Identification and characterization of a novel rat ov-serpin family member, trespin. J Biol Chem. 2002b;277:26412–21. - PubMed
-
- Danielpour D. Functions and regulation of transforming growth factor-beta (TGF-beta) in the prostate. Eur J Cancer. 2005;41:846–57. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
