

Up-regulating sphingosine 1-phosphate receptor-2 signaling impairs chemotactic, wound-healing, and morphogenetic responses in senescent endothelial cells
- PMID: 18765664
- PMCID: PMC2573088
- DOI: 10.1074/jbc.M804392200
Up-regulating sphingosine 1-phosphate receptor-2 signaling impairs chemotactic, wound-healing, and morphogenetic responses in senescent endothelial cells
Abstract
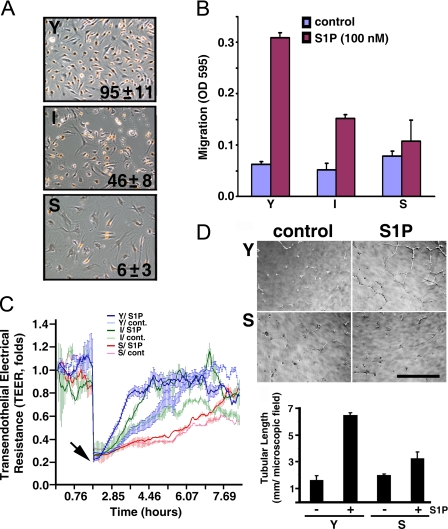
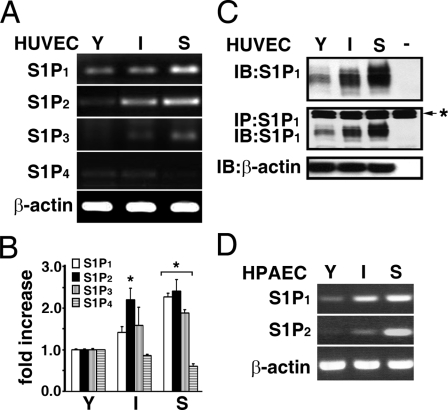
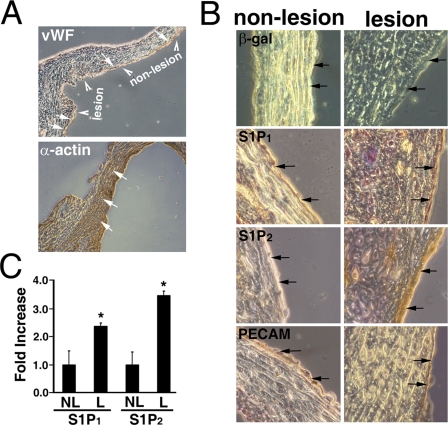
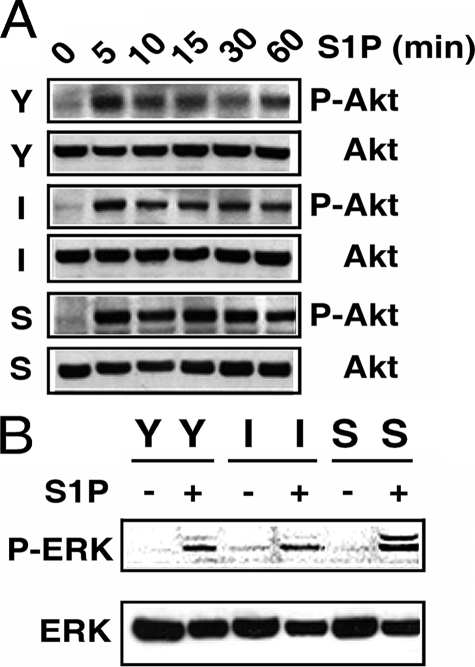
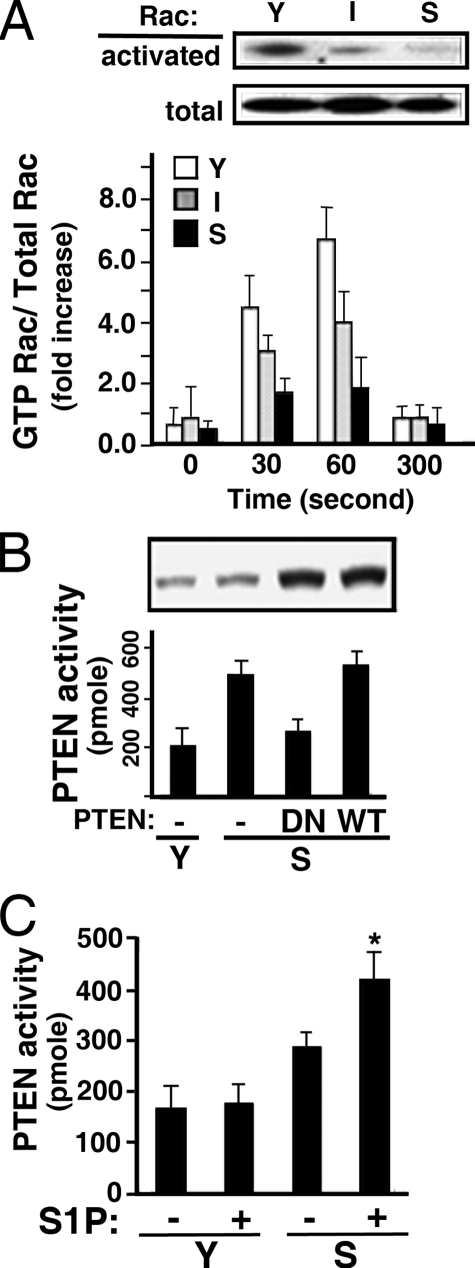
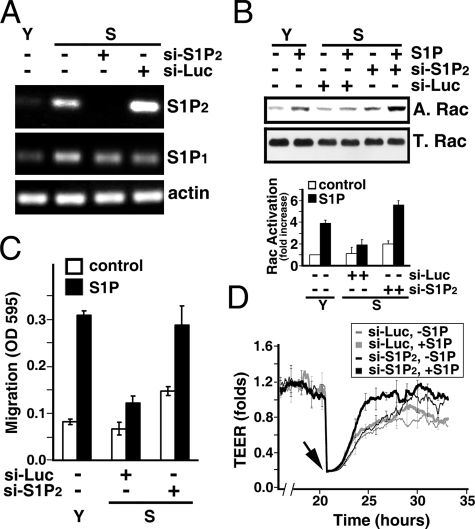
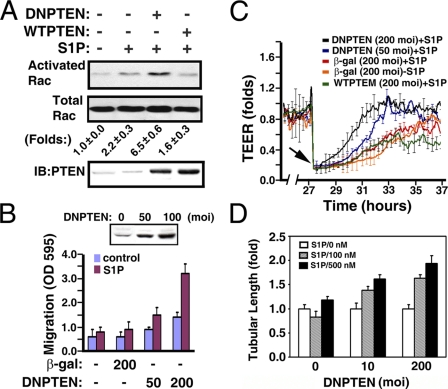
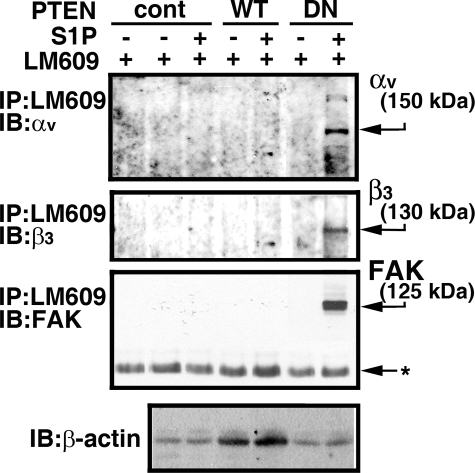
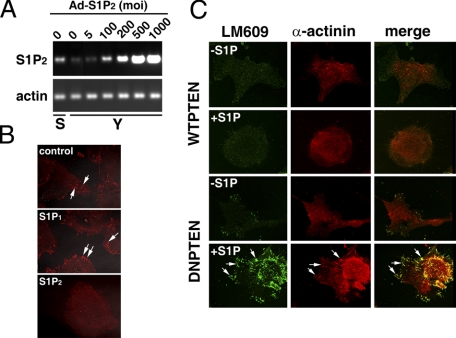
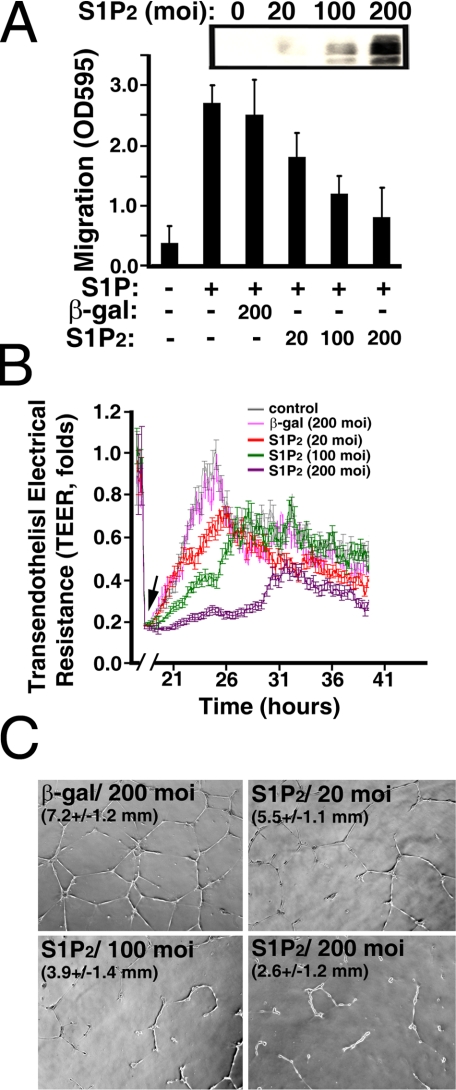
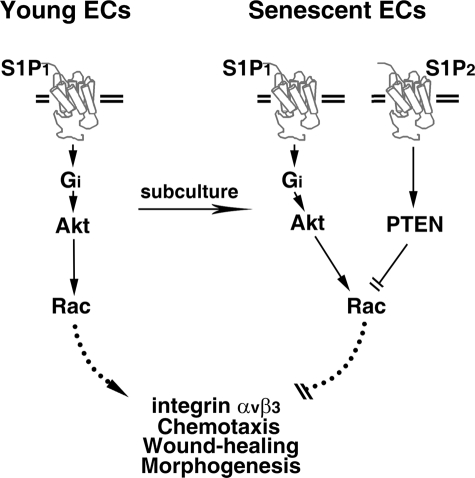
Vascular endothelial cells (ECs) have a finite lifespan when cultured in vitro and eventually enter an irreversible growth arrest state called "cellular senescence." It has been shown that sphingolipids may be involved in senescence; however, the molecular links involved are poorly understood. In this study, we investigated the signaling and functions of sphingosine 1-phosphate (S1P), a serum-borne bioactive sphingolipid, in ECs of different in vitro ages. We observed that S1P-regulated responses are significantly inhibited and the S1P(1-3) receptor subtypes are markedly increased in senescent ECs. Increased expression of S1P(1) and S1P(2) was also observed in the lesion regions of atherosclerotic endothelium, where senescent ECs have been identified in vivo. S1P-induced Akt and ERK1/2 activation were comparable between ECs of different in vitro ages; however, PTEN (phosphatase and tensin homolog deleted on chromosome 10) activity was significantly elevated and Rac activation was inhibited in senescent ECs. Rac activation and senescent-associated impairments were restored in senescent ECs by the expression of dominant-negative PTEN and by knocking down S1P(2) receptors. Furthermore, the senescent-associated impairments were induced in young ECs by the expression of S1P(2) to a level similar to that of in vitro senescence. These results indicate that the impairment of function in senescent ECs in culture is mediated by an increase in S1P signaling through S1P(2)-mediated activation of the lipid phosphatase PTEN.
Figures
References
-
- Moolenaar, W. H. (1999) Exp. Cell Res. 253 230-238 - PubMed
-
- Spiegel, S. (1999) J. Leukoc. Biol. 65 341-344 - PubMed
-
- Igarashi, Y., and Yatomi, Y. (1998) Acta Biochim. Pol. 45 299-309 - PubMed
-
- Hla, T., Lee, M. J., Ancellin, N., Liu, C. H., Thangada, S., Thompson, B. D., and Kluk, M. (1999) Biochem. Pharmacol. 58 201-207 - PubMed
-
- An, S., Goetzl, E. J., and Lee, H. (1998) J. Cell. Biochem. Suppl. 30-31 147-157 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
