

The gamma interferon (IFN-gamma)-inducible GTP-binding protein IGTP is necessary for toxoplasma vacuolar disruption and induces parasite egression in IFN-gamma-stimulated astrocytes
- PMID: 18765738
- PMCID: PMC2573374
- DOI: 10.1128/IAI.01288-07
The gamma interferon (IFN-gamma)-inducible GTP-binding protein IGTP is necessary for toxoplasma vacuolar disruption and induces parasite egression in IFN-gamma-stimulated astrocytes
Abstract
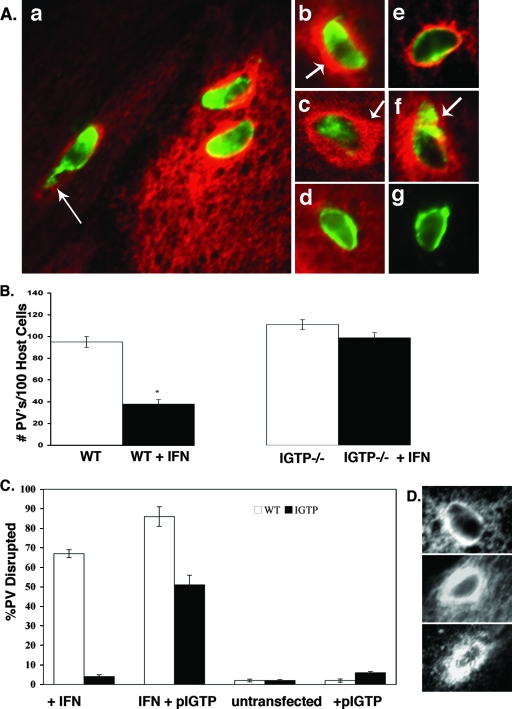
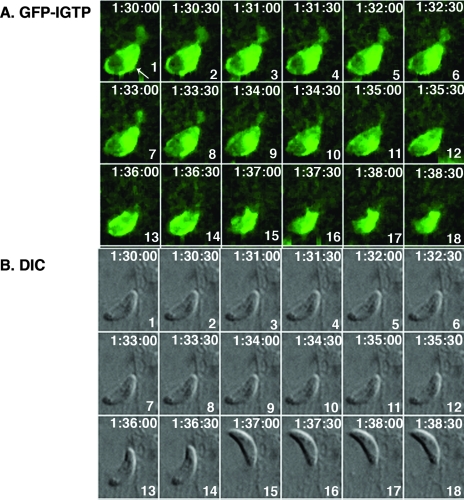
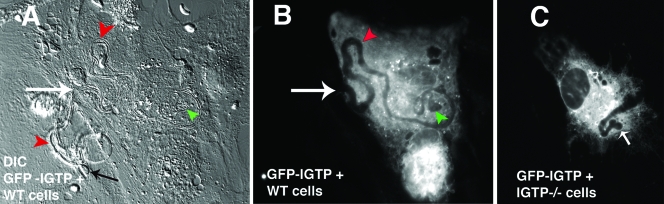
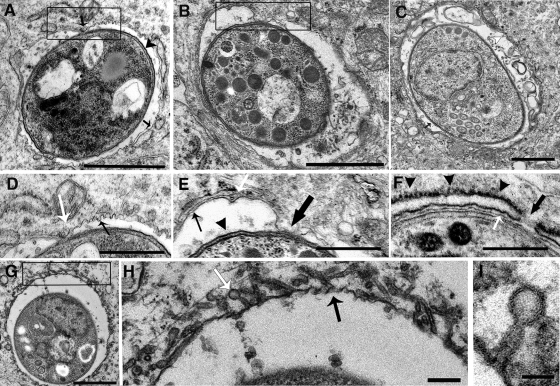
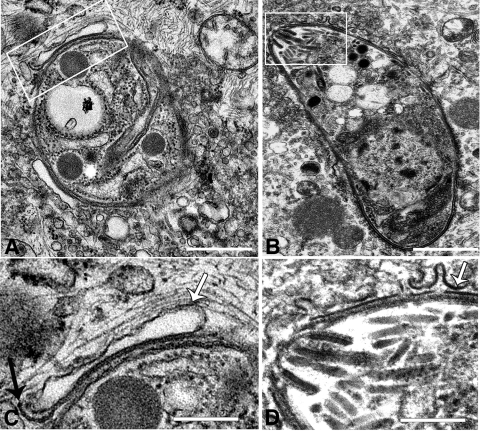
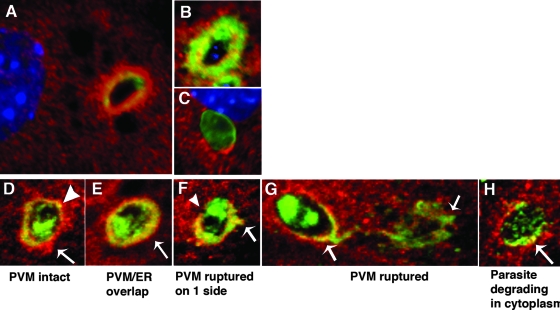
Toxoplasma gondii is a common central nervous system infection in individuals with immunocompromised immune systems, such as AIDS patients. Gamma interferon (IFN-gamma) is the main cytokine mediating protection against T. gondii. Our previous studies found that IFN-gamma significantly inhibits T. gondii in astrocytes via an IFN-gamma-inducible GTP-binding protein (IGTP)-dependent mechanism. The IGTP-dependent-, IFN-gamma-stimulated inhibition is not understood, but recent studies found that IGTP induces disruption of the parasitophorous vacuole (PV) in macrophages. In the current study, we have further investigated the mechanism of IFN-gamma inhibition and the role of IGTP in the vacuolar disruption in murine astrocytes. Vacuolar disruption was found to be dependent upon IGTP, as PV disruption was not observed in IGTP-deficient (IGTP(-/-)) astrocytes and PV disruption could be induced in IGTP(-/-) astrocytes transfected with IGTP. Live-cell imaging studies using green fluorescent protein-IGTP found that IGTP is delivered to the PV via the host cell endoplasmic reticulum (ER) early after invasion and that IGTP condenses into vesicle-like structures on the vacuole just prior to PV disruption, suggesting that IGTP is involved in PV disruption. Intravacuolar movement of the parasite occurred just prior to PV disruption. In some instances, IFN-gamma induced parasite egression. Electron microscopy and immunofluorescence studies indicate that the host cell ER fuses with the PV prior to vacuolar disruption. On the basis of these results, we postulate a mechanism by which ER/PV fusion is a crucial event in PV disruption. Fusion of the ER with the PV, releasing calcium into the vacuole, may also be the mechanism by which intravacuolar parasite movement and IFN-gamma-induced parasite egression occur.
Figures

Similar articles
-
Gamma interferon-induced inhibition of Toxoplasma gondii in astrocytes is mediated by IGTP.Infect Immun. 2001 Sep;69(9):5573-6. doi: 10.1128/IAI.69.9.5573-5576.2001. Infect Immun. 2001. PMID: 11500431 Free PMC article.
-
The function of gamma interferon-inducible GTP-binding protein IGTP in host resistance to Toxoplasma gondii is Stat1 dependent and requires expression in both hematopoietic and nonhematopoietic cellular compartments.Infect Immun. 2002 Dec;70(12):6933-9. doi: 10.1128/IAI.70.12.6933-6939.2002. Infect Immun. 2002. PMID: 12438372 Free PMC article.
-
Disruption of Toxoplasma gondii parasitophorous vacuoles by the mouse p47-resistance GTPases.PLoS Pathog. 2005 Nov;1(3):e24. doi: 10.1371/journal.ppat.0010024. Epub 2005 Nov 18. PLoS Pathog. 2005. PMID: 16304607 Free PMC article.
-
Biogenesis of and activities at the Toxoplasma gondii parasitophorous vacuole membrane.Subcell Biochem. 2008;47:155-64. doi: 10.1007/978-0-387-78267-6_12. Subcell Biochem. 2008. PMID: 18512349 Review.
-
Toxoplasma gondii and the Immunity-Related GTPase (IRG) resistance system in mice: a review.Mem Inst Oswaldo Cruz. 2009 Mar;104(2):234-40. doi: 10.1590/s0074-02762009000200016. Mem Inst Oswaldo Cruz. 2009. PMID: 19430648 Review.
Cited by
-
Molecular mechanism for the control of virulent Toxoplasma gondii infections in wild-derived mice.Nat Commun. 2019 Mar 15;10(1):1233. doi: 10.1038/s41467-019-09200-2. Nat Commun. 2019. PMID: 30874554 Free PMC article.
-
Guanylate binding proteins directly attack Toxoplasma gondii via supramolecular complexes.Elife. 2016 Jan 27;5:e11479. doi: 10.7554/eLife.11479. Elife. 2016. PMID: 26814575 Free PMC article.
-
Interferon-γ restricts Toxoplasma gondii development in murine skeletal muscle cells via nitric oxide production and immunity-related GTPases.PLoS One. 2012;7(9):e45440. doi: 10.1371/journal.pone.0045440. Epub 2012 Sep 14. PLoS One. 2012. PMID: 23024821 Free PMC article.
-
A dedicated promoter drives constitutive expression of the cell-autonomous immune resistance GTPase, Irga6 (IIGP1) in mouse liver.PLoS One. 2009 Aug 26;4(8):e6787. doi: 10.1371/journal.pone.0006787. PLoS One. 2009. PMID: 20368812 Free PMC article.
-
Identification of a novel protein complex essential for effector translocation across the parasitophorous vacuole membrane of Toxoplasma gondii.PLoS Pathog. 2018 Jan 22;14(1):e1006828. doi: 10.1371/journal.ppat.1006828. eCollection 2018 Jan. PLoS Pathog. 2018. PMID: 29357375 Free PMC article.
References
-
- Adams, L. B., J. B. Hibbs, Jr., R. R. Taintor, and J. L. Krahenbuhl. 1990. Microbiostatic effect of murine-activated macrophages for Toxoplasma gondii. Role for synthesis of inorganic nitrogen oxides from L-arginine. J. Immunol. 1442725-2729. - PubMed
-
- Arrizabalaga, G., and J. C. Boothroyd. 2004. Role of calcium during Toxoplasma gondii invasion and egress. Int. J. Parasitol. 34361-368. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
