

Cutting edge: Th1 cells facilitate the entry of Th17 cells to the central nervous system during experimental autoimmune encephalomyelitis
- PMID: 18768826
- PMCID: PMC2619513
- DOI: 10.4049/jimmunol.181.6.3750
Cutting edge: Th1 cells facilitate the entry of Th17 cells to the central nervous system during experimental autoimmune encephalomyelitis
Abstract
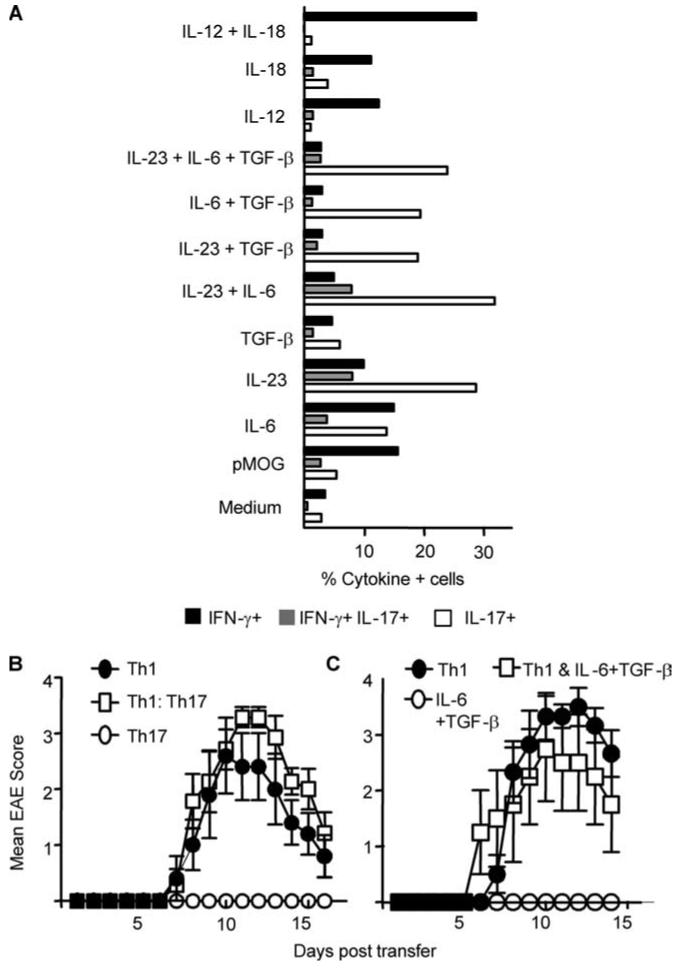
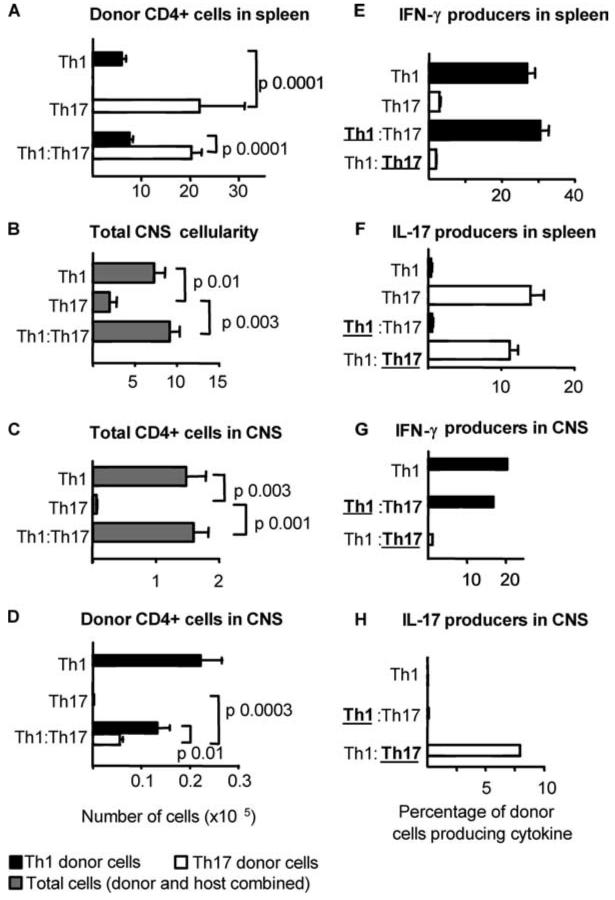
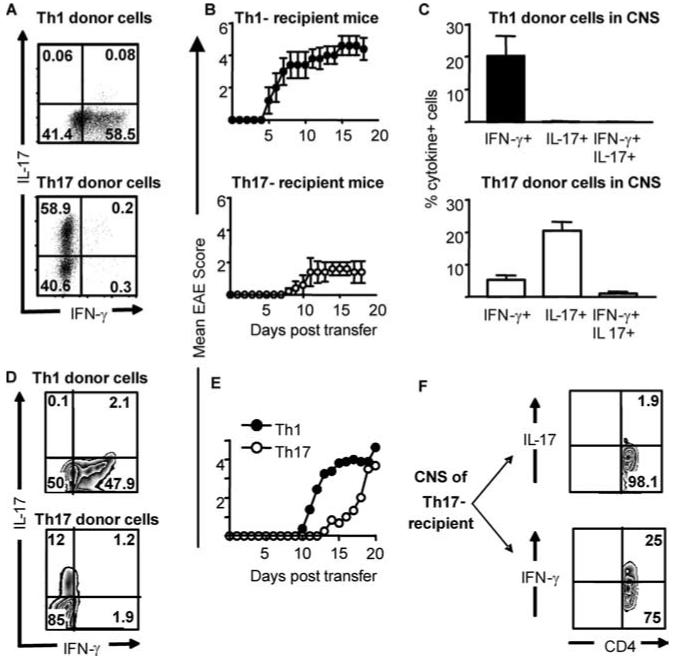
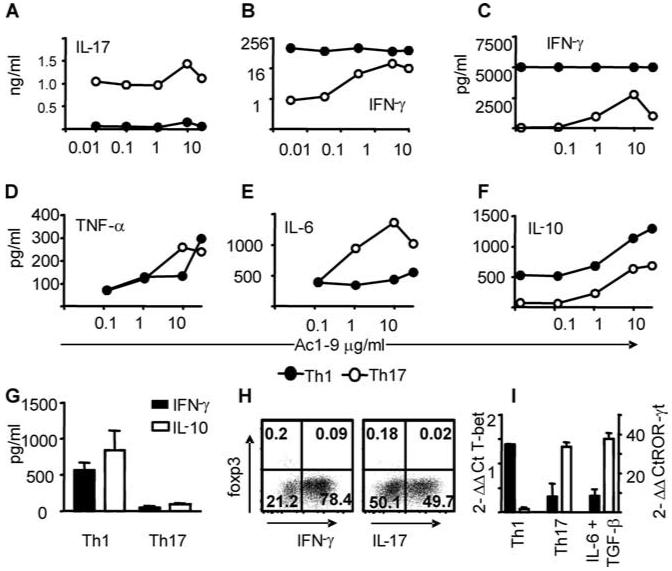
It has recently been proposed that experimental autoimmune encephalomyelitis, once considered the classical Th1 disease, is predominantly Th17 driven. In this study we show that myelin-reactive Th1 preparations devoid of contaminating IL-17(+) cells are highly pathogenic. In contrast, Th17 preparations lacking IFN-gamma(+) cells do not cause disease. Our key observation is that only Th1 cells can access the noninflamed CNS. Once Th1 cells establish the experimental autoimmune encephalomyelitis lesion, Th17 cells appear in the CNS. These data shed important new light on the ability of Th1 vs Th17 cells to access inflamed vs normal tissue. Because the IL-17-triggered release of chemokines by stromal cells could attract many other immune cells, allowing Th17 cells to access the tissues only under conditions of inflammation may be a key process limiting (auto)immune pathology. This has major implications for the design of therapeutic interventions, many of which are now aiming at Th17 rather than Th1 cells.
Figures
References
-
- Cua DJ, Sherlock J, Chen Y, Murphy CA, Joyce B, Seymour B, Lucian L, To W, Kwan S, Churakova T, et al. Interleukin-23 rather than interleukin-12 is the critical cytokine for autoimmune inflammation of the brain. Nature. 2003;421:744–748. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
