

Functional analysis of the cellulose synthase-like genes CSLD1, CSLD2, and CSLD4 in tip-growing Arabidopsis cells
- PMID: 18768911
- PMCID: PMC2577265
- DOI: 10.1104/pp.108.121939
Functional analysis of the cellulose synthase-like genes CSLD1, CSLD2, and CSLD4 in tip-growing Arabidopsis cells
Abstract
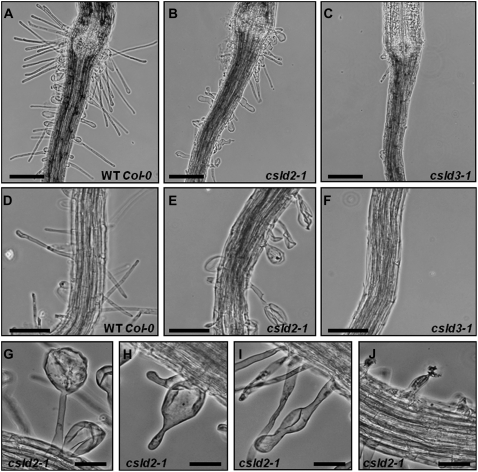
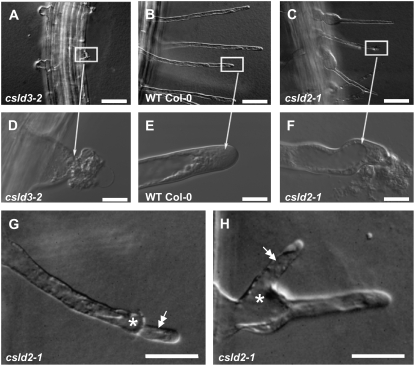
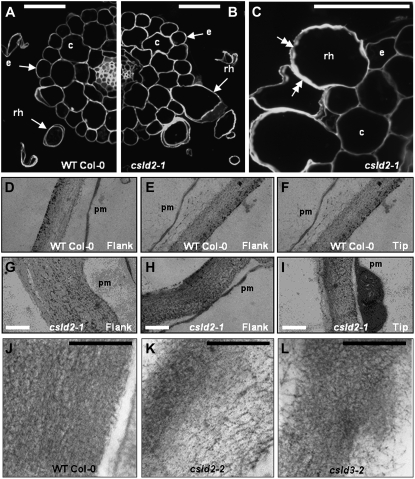
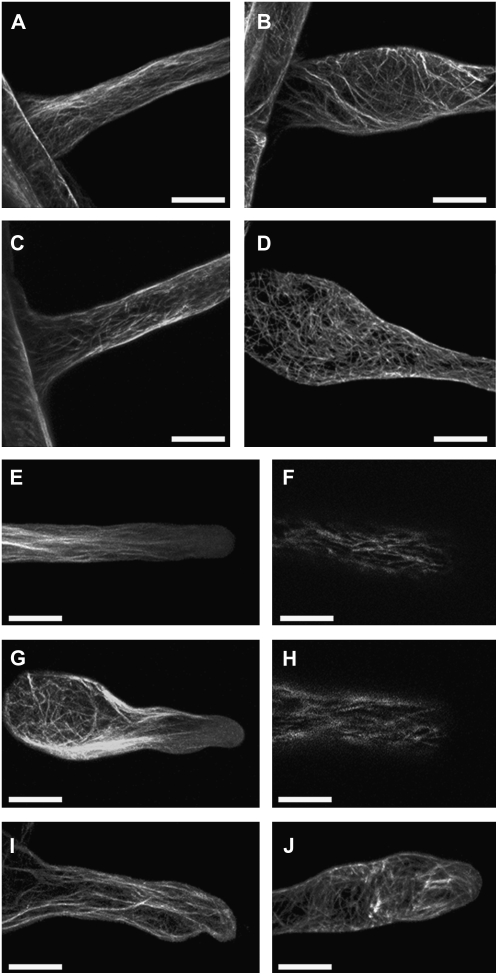
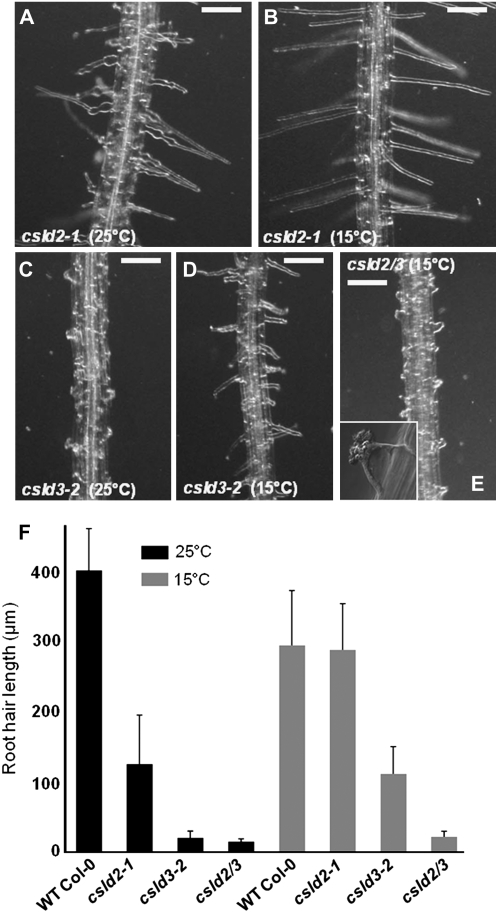
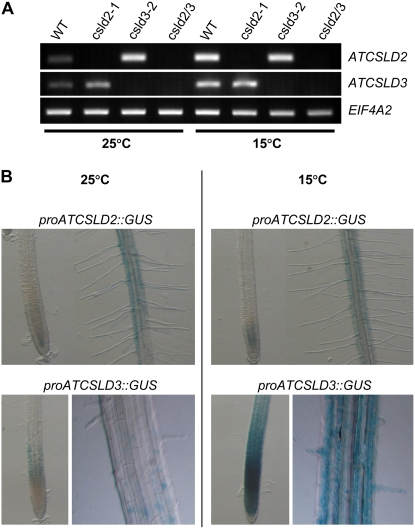
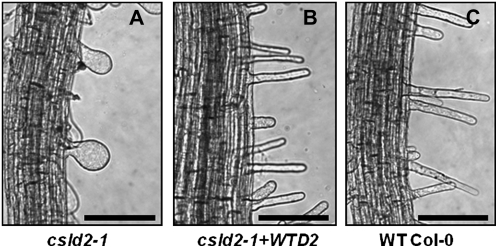
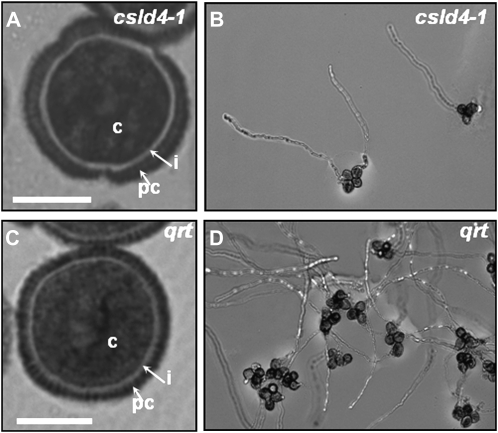
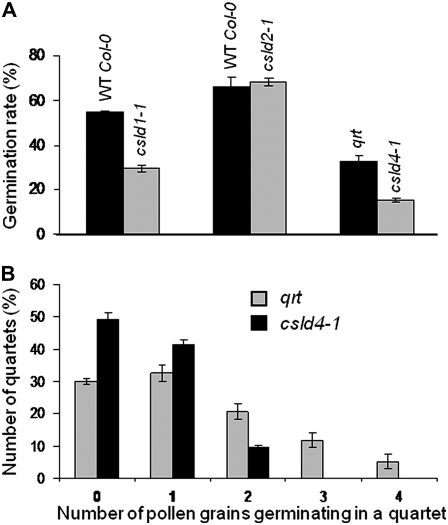
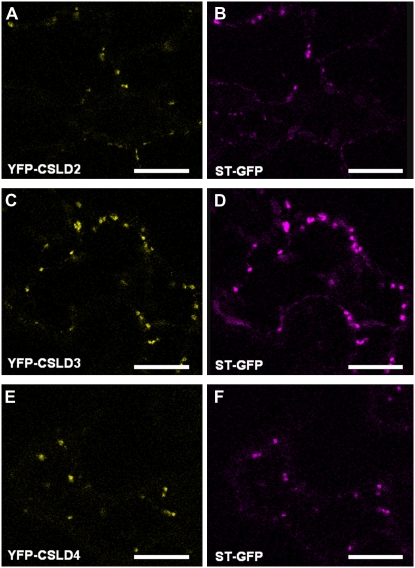
A reverse genetic approach was used to investigate the functions of three members of the cellulose synthase superfamily in Arabidopsis (Arabidopsis thaliana), CELLULOSE SYNTHASE-LIKE D1 (CSLD1), CSLD2, and CSLD4. CSLD2 is required for normal root hair growth but has a different role from that previously described for CSLD3 (KOJAK). CSLD2 is required during a later stage of hair development than CSLD3, and CSLD2 mutants produce root hairs with a range of abnormalities, with many root hairs rupturing late in development. Remarkably, though, it was often the case that in CSLD2 mutants, tip growth would resume after rupturing of root hairs. In silico, semiquantitative reverse transcription-polymerase chain reaction, and promoter-reporter construct analyses indicated that the expression of both CSLD2 and CSLD3 is elevated at reduced temperatures, and the phenotypes of mutants homozygous for insertions in these genes were partially rescued by reduced temperature growth. However, this was not the case for a double mutant homozygous for insertions in both CSLD2 and CSLD3, suggesting that there may be partial redundancy in the functions of these genes. Mutants in CSLD1 and CSLD4 had a defect in male transmission, and plants heterozygous for insertions in CSLD1 or CSLD4 were defective in their ability to produce pollen tubes, although the number and morphology of pollen grains was normal. We propose that the CSLD family of putative glycosyltransferases synthesize a polysaccharide that has a specialized structural role in the cell walls of tip-growing cells.
Figures
Similar articles
-
The cooperative activities of CSLD2, CSLD3, and CSLD5 are required for normal Arabidopsis development.Mol Plant. 2011 Nov;4(6):1024-37. doi: 10.1093/mp/ssr026. Epub 2011 Apr 6. Mol Plant. 2011. PMID: 21471331
-
Arabidopsis CSLD1 and CSLD4 are required for cellulose deposition and normal growth of pollen tubes.J Exp Bot. 2011 Oct;62(14):5161-77. doi: 10.1093/jxb/err221. Epub 2011 Jul 15. J Exp Bot. 2011. PMID: 21765162 Free PMC article.
-
Divergence and Redundancy in CSLD2 and CSLD3 Function During Arabidopsis Thaliana Root Hair and Female Gametophyte Development.Front Plant Sci. 2012 May 29;3:111. doi: 10.3389/fpls.2012.00111. eCollection 2012. Front Plant Sci. 2012. PMID: 22661983 Free PMC article.
-
Root hair-specific disruption of cellulose and xyloglucan in AtCSLD3 mutants, and factors affecting the post-rupture resumption of mutant root hair growth.Planta. 2011 May;233(5):985-99. doi: 10.1007/s00425-011-1355-6. Epub 2011 Jan 29. Planta. 2011. PMID: 21279381
-
Building a hair: tip growth in Arabidopsis thaliana root hairs.Philos Trans R Soc Lond B Biol Sci. 2002 Jun 29;357(1422):815-21. doi: 10.1098/rstb.2002.1092. Philos Trans R Soc Lond B Biol Sci. 2002. PMID: 12079677 Free PMC article. Review.
Cited by
-
Arabidopsis CSLD5 Functions in Cell Plate Formation in a Cell Cycle-Dependent Manner.Plant Cell. 2016 Jul;28(7):1722-37. doi: 10.1105/tpc.16.00203. Epub 2016 Jun 27. Plant Cell. 2016. PMID: 27354558 Free PMC article.
-
High throughput generation of promoter reporter (GFP) transgenic lines of low expressing genes in Arabidopsis and analysis of their expression patterns.Plant Methods. 2010 Aug 6;6:18. doi: 10.1186/1746-4811-6-18. Plant Methods. 2010. PMID: 20687964 Free PMC article.
-
Revised Phylogeny of the Cellulose Synthase Gene Superfamily: Insights into Cell Wall Evolution.Plant Physiol. 2018 Jul;177(3):1124-1141. doi: 10.1104/pp.17.01718. Epub 2018 May 20. Plant Physiol. 2018. PMID: 29780036 Free PMC article.
-
Evolution, gene expression profiling and 3D modeling of CSLD proteins in cotton.BMC Plant Biol. 2017 Jul 10;17(1):119. doi: 10.1186/s12870-017-1063-x. BMC Plant Biol. 2017. PMID: 28693426 Free PMC article.
-
Xyloglucan deficiency leads to a reduction in turgor pressure and changes in cell wall properties, affecting early seedling establishment.Curr Biol. 2024 May 20;34(10):2094-2106.e6. doi: 10.1016/j.cub.2024.04.016. Epub 2024 Apr 26. Curr Biol. 2024. PMID: 38677280 Free PMC article.
References
-
- Bacic A, Harris PJ, Stone BA (1988) Structure and function of plant cell walls. In J Preiss, ed, The Biochemistry of Plants, Vol 14. Academic Press, New York, pp 297–371
-
- Bernal AJ, Jensen JK, Harholt J, Sørensen S, Moller I, Blaukopf C, Johansen B, de Lotto R, Pauly M, Schelle HV, et al (2007) Disruption of ATCSLD5 results in reduced growth, reduced xylan and homogalacturonan synthase activity and altered xylan occurrence in Arabidopsis. Plant J 52 791–802 - PubMed
-
- Blancaflor EB, Hasenstein KH (1995) Growth and microtubule orientation of Zea mays roots subjected to osmotic stress. Int J Plant Sci 156 774–783 - PubMed
-
- Blancaflor EB, Hou G, Chapman KD (2003) Elevated levels of N-lauroethanolamine, an endogenous constituent of desiccated seeds, disrupt normal root development in Arabidopsis thaliana seedlings. Planta 217 206–217 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
