

Molecular and transmission characteristics of primary-passaged ovine scrapie isolates in conventional and ovine PrP transgenic mice
- PMID: 18768980
- PMCID: PMC2573291
- DOI: 10.1128/JVI.01454-08
Molecular and transmission characteristics of primary-passaged ovine scrapie isolates in conventional and ovine PrP transgenic mice
Abstract
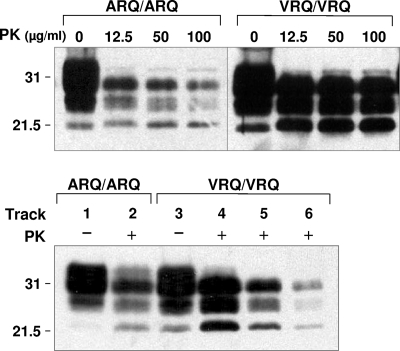
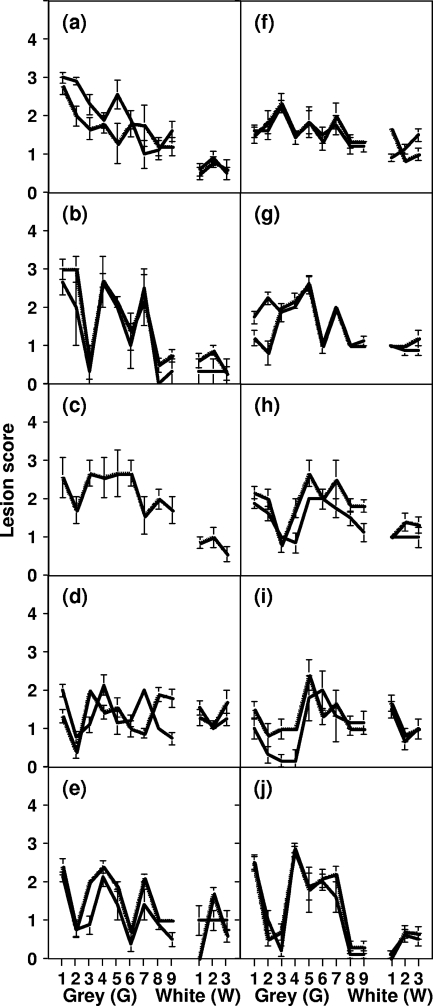
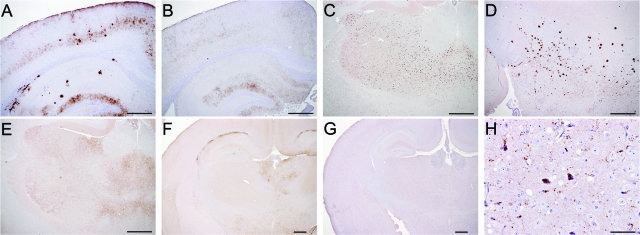
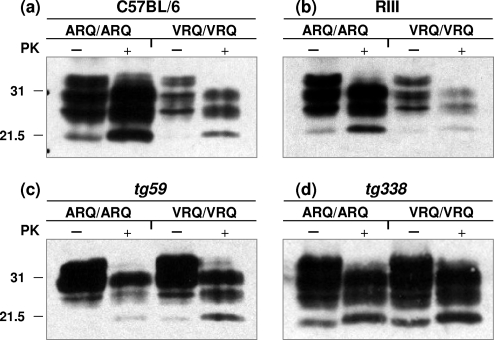
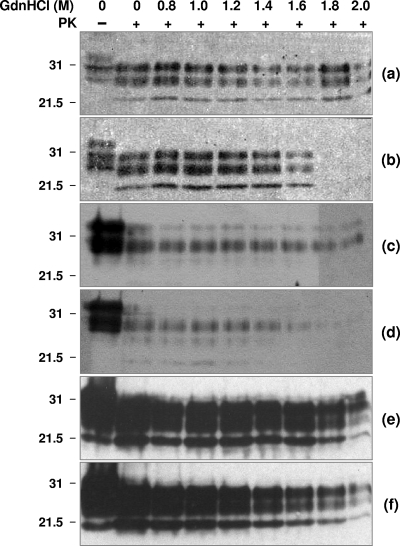
A more complete assessment of ovine prion strain diversity will be achieved by complementing biological strain typing in conventional and ovine PrP transgenic mice with a biochemical analysis of the resultant PrPSc. This will provide a correlation between ovine prion strain phenotype and the molecular nature of different PrP conformers associated with particular prion strains. Here, we have compared the molecular and transmission characteristics of ovine ARQ/ARQ and VRQ/VRQ scrapie isolates following primary passage in tg338 (VRQ) and tg59 (ARQ) ovine PrP transgenic mice and the conventional mouse lines C57BL/6 (Prnp(a)), RIII (Prnp(a)), and VM (Prnp(b)). Our data show that these different genotypes of scrapie isolates display similar incubation periods of >350 days in conventional and tg59 mice. Facilitated transmission of sheep scrapie isolates occurred in tg338 mice, with incubation times reduced to 64 days for VRQ/VRQ inocula and to </=210 days for ARQ/ARQ samples. Distinct genotype-specific lesion profiles were seen in the brains of conventional and tg59 mice with prion disease, which was accompanied by the accumulation of more conformationally stable PrPSc, following inoculation with ARQ/ARQ compared to VRQ/VRQ scrapie isolates. In contrast, the lesion profiles, quantities, and stability of PrPSc induced by the same inocula in tg338 mice were more similar than in the other mouse lines. Our data show that primary transmission of different genotypes of ovine prions is associated with the formation of different conformers of PrPSc with distinct molecular properties and provide the basis of a molecular approach to identify the true diversity of ovine prion strains.
Figures
Similar articles
-
Propagation of ovine prions from "poor" transmitter scrapie isolates in ovine PrP transgenic mice.Exp Mol Pathol. 2012 Feb;92(1):167-74. doi: 10.1016/j.yexmp.2011.11.004. Epub 2011 Nov 19. Exp Mol Pathol. 2012. PMID: 22120785
-
In vitro amplification of ovine prions from scrapie-infected sheep from Great Britain reveals distinct patterns of propagation.BMC Vet Res. 2012 Nov 15;8:223. doi: 10.1186/1746-6148-8-223. BMC Vet Res. 2012. PMID: 23153009 Free PMC article.
-
Strain Typing of Classical Scrapie and Bovine Spongiform Encephalopathy (BSE) by Using Ovine PrP (ARQ/ARQ) Overexpressing Transgenic Mice.Int J Mol Sci. 2022 Jun 16;23(12):6744. doi: 10.3390/ijms23126744. Int J Mol Sci. 2022. PMID: 35743187 Free PMC article.
-
Prion encephalopathies of animals and humans.Dev Biol Stand. 1993;80:31-44. Dev Biol Stand. 1993. PMID: 8270114 Review.
-
Genetic and infectious prion diseases.Arch Neurol. 1993 Nov;50(11):1129-53. doi: 10.1001/archneur.1993.00540110011002. Arch Neurol. 1993. PMID: 8105771 Review.
Cited by
-
Evolutionary biology and the risk of scrapie disease in sheep.Open Vet J. 2018;8(3):282-294. doi: 10.4314/ovj.v8i3.7. Epub 2018 Aug 7. Open Vet J. 2018. PMID: 30148080 Free PMC article. Review.
-
Use of murine bioassay to resolve ovine transmissible spongiform encephalopathy cases showing a bovine spongiform encephalopathy molecular profile.Brain Pathol. 2012 May;22(3):265-79. doi: 10.1111/j.1750-3639.2011.00526.x. Epub 2011 Oct 13. Brain Pathol. 2012. PMID: 21919992 Free PMC article.
-
Genetic modulation of CWD prion propagation in cervid PrP Drosophila.Biochem J. 2023 Oct 11;480(19):1485-1501. doi: 10.1042/BCJ20230247. Biochem J. 2023. PMID: 37747806 Free PMC article.
-
Evidence of p75 Neurotrophin Receptor Involvement in the Central Nervous System Pathogenesis of Classical Scrapie in Sheep and a Transgenic Mouse Model.Int J Mol Sci. 2021 Mar 8;22(5):2714. doi: 10.3390/ijms22052714. Int J Mol Sci. 2021. PMID: 33800240 Free PMC article.
-
Mammalian prion propagation in PrP transgenic Drosophila.Brain. 2018 Sep 1;141(9):2700-2710. doi: 10.1093/brain/awy183. Brain. 2018. PMID: 29985975 Free PMC article.
References
-
- Aguzzi, A., M. Heikenwalder, and M. Polymenidou. 2007. Insights into prion strains and neurotoxicity. Nat. Rev. Mol. Cell Biol. 8552-561. - PubMed
-
- Anderson, M., O. V. Bocharova, N. Makarava, L. Breydo, V. V. Salnikov, and I. V. Baskakov. 2006. Polymorphism and ultrastructural organization of prion protein amyloid fibrils: an insight from high resolution atomic force microscopy. J. Mol. Biol. 358580-596. - PubMed
-
- The Animals (Scientific Procedures) Act. 1986. Eliz. 2, chapter 14. Her Majesty's Stationery Office, London, United Kingdom.
-
- Asante, E. A., J. M. Linehan, M. Desbruslais, S. Joiner, I. Gowland, A. L. Wood, J. Welch, A. F. Hill, S. E. Lloyd, J. D. Wadsworth, and J. Collinge. 2002. BSE prions propagate as either variant CJD-like or sporadic CJD-like prion strains in transgenic mice expressing human prion protein. EMBO J. 216358-6366. - PMC - PubMed
-
- Baron, T., C. Crozet, A. G. Biacabe, S. Philippe, J. Verchere, A. Bencsik, J. Y. Madec, D. Calavas, and J. Samarut. 2004. Molecular analysis of the protease-resistant prion protein in scrapie and bovine spongiform encephalopathy transmitted to ovine transgenic and wild-type mice. J. Virol. 786243-6251. - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
