

The insulin paradox: aging, proteotoxicity and neurodegeneration
- PMID: 18769445
- PMCID: PMC2692886
- DOI: 10.1038/nrn2474
The insulin paradox: aging, proteotoxicity and neurodegeneration
Abstract
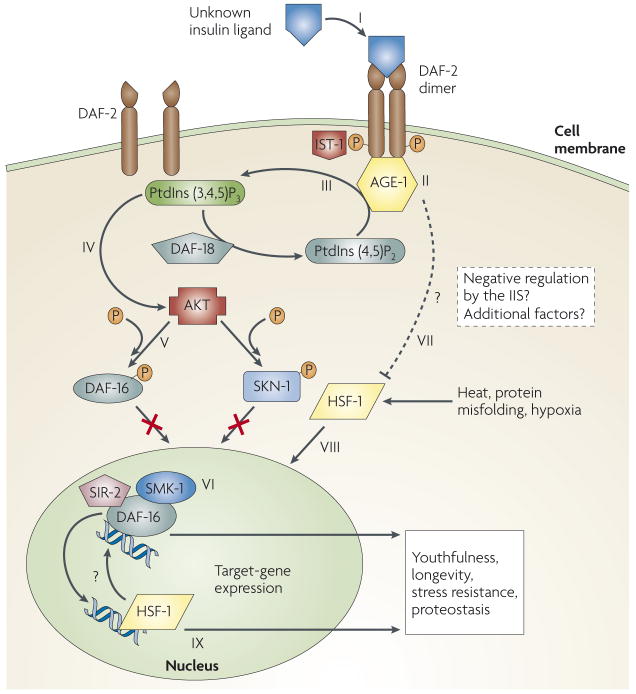
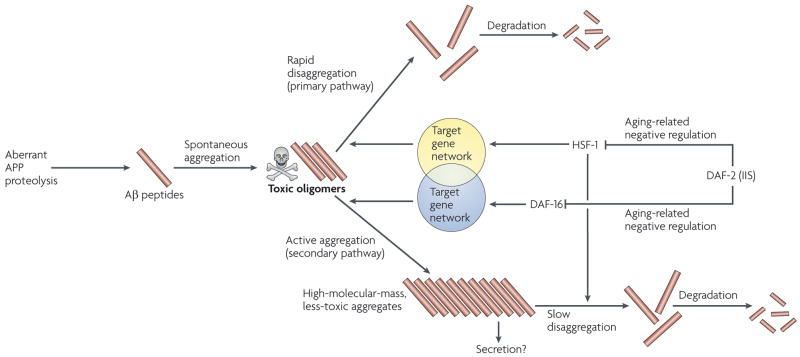
Distinct human neurodegenerative diseases share remarkably similar temporal emergence patterns, even though different toxic proteins are involved in their onset. Typically, familial neurodegenerative diseases emerge during the fifth decade of life, whereas sporadic cases do not exhibit symptoms earlier than the seventh decade. Recently, mechanistic links between the aging process and toxic protein aggregation, a common hallmark of neurodegenerative diseases, have been revealed. The insulin/insulin-like growth factor 1 (IGF1) signalling pathway - a lifespan, metabolism and stress-resistance regulator - links neurodegeneration to the aging process. Thus, although a reduction of insulin signalling can result in diabetes, its reduction can also increase longevity and delay the onset of protein-aggregation-mediated toxicity. Here we review this apparent paradox and delineate the therapeutic potential of manipulating the insulin/IGF1 signalling pathway for the treatment of neurodegenerative diseases.
Figures


References
-
- Reichel W. The biology of aging. J Am Geriatr Soc. 1966;14:431–436. - PubMed
-
- Bishop NA, Guarente L. Genetic links between diet and lifespan: shared mechanisms from yeast to humans. Nature Rev Genet. 2007;8:835–844. - PubMed
-
- Mair W, Dillin A. Aging and survival: the genetics of life span extension by dietary restriction. Annu Rev Biochem. 2008;77:727–754. - PubMed
-
- Kenyon C, Chang J, Gensch E, Rudner A, Tabtiang R. A C. elegans mutant that lives twice as long as wild type. Nature. 1993;366:461–464. - PubMed
-
- Giannakou ME, Partridge L. Role of insulin-like signalling in Drosophila lifespan. Trends Biochem Sci. 2007;32:180–188. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
